
Quel est le rôle de la socialité dans la démographie du zèbre des plaines Equus quagga en contexte de prédation ?
Camille Vitet
Email: camille.vitet@cefe.cnrs.fr
(Version PDF)
Résumé
Dans le cadre de mon projet de terrain, je me suis rendue au parc national de Matopos. Ce parc n’abrite pas de prédateurs importants pour les zèbres : lions, lycaons sont absents, et la présence des hyènes tachetées est anecdotique. Le but était de collecter des données de tailles de groupe et de taux de recrutement (nombre de jeunes de moins d’un an) pour la population de zèbre des plaines occupant cette zone, et également de prendre des photos des individus pour être capable de les identifier de nouveau un an après à partir du motif de leurs rayures et ainsi pouvoir estimer leur survie. L’objectif était ensuite de comparer ces données avec celles des populations de zèbres du parc national de Hwange, particulièrement celle de la zone de Main Camp où le taux de prédation est élevé pour les zèbres (2.7 lions/100 km²)1 afin de mieux comprendre l’impact d’une contrainte environnementale telle que le risque de prédation sur la démographie de cette espèce.
Au cours de la visite exploratoire du parc de Matopos, aucune donnée n’a pu être collectée puisqu’aucun zèbre n’a été vu, en grande partie dû à la végétation trop dense en cette saison qui réduit considérablement la visibilité. Par conséquent, j’ai choisi de réaliser des prospections dans d’autres zones du pays, à savoir le parc national de Kazuma Pan, le parc national de Zambezi et la zone ouest du parc national de Hwange. Alors qu’aucun zèbre n’a été vu dans le parc national de Kazuma Pan, 3 groupes ont été observés dans le parc national de Zambezi mais la taille de groupe et le recrutement pour 2 d’entre eux n’ont pu être déterminé avec exactitude à cause de la végétation trop dense. Par conséquent, pour des raisons de visibilité, j’ai choisi de me concentrer uniquement sur la zone ouest du parc national de Hwange qui est celle qui offre les meilleures conditions pour observer les zèbres. En 5 jours sur place, j’y ai observé 4 groupes. Cette taille d’échantillon trop petite ne permet aucune analyse robuste. Néanmoins, les résultats suggèrent que les tailles de groupes sont similaires entre les groupes de Main Camp et ceux de la zone ouest. Par contre, il semble que le taux de recrutement (proportion de poulains dans les groupes) soit plus faible pour les zèbres de la zone ouest.
Justification
Le fait de vivre en groupe est généralement considéré comme une protection contre les prédateurs, en raison des effets de dilution du risque et de la vigilance collective qu’un groupe apporte 2,3. Une partie de cette sécurité accrue est généralement échangée contre une vigilance individuelle réduite 4 conduisant à une augmentation de l’approvisionnement en ressource. Les individus appartenant à des groupes plus grands pourraient également utiliser des zones plus profitables mais plus risquées mais cela a été rarement testé. En règle générale, nous nous attendons à ce que la taille du groupe améliore la fitness des individus en raison de la réduction du risque et/ou de l’augmentation de l’approvisionnement, jusqu’à ce que les coûts d’une compétition accrue à l’intérieur du groupe en compensent les avantages 5,6. Cependant, peu d’études ont été en mesure de tester directement la relation entre la performance démographique des individus (survie, succès reproducteur, croissance) et la taille des groupes. Cela s’explique par le fait que très peu d’études démographiques existent sur des espèces vivant dans des groupes assez stables ou bien qu’elles se soient limitées à quelques groupes ou aient été conduites en l’absence de prédation. Ainsi, le lien entre effets de dilution/détection et performances démographiques dans un contexte de risque de prédation reste essentiellement théorique, et les rares études réalisées ne soutiennent pas toujours la théorie7.
Dans ce contexte, l’un des objectifs de ma thèse est (1) d’étudier l’effet de la taille de groupe sur la fitness des individus en déterminant les coûts et les bénéfices liés à la taille du groupe en termes de vigilance, d’interactions agonistiques et d’approvisionnement en ressources, puis (2) d’évaluer si ces coûts et bénéfices se traduisent par des signaux démographiques.
Mon étude utilise le zèbre des plaines Equus quagga comme modèle et la Zone Atelier CNRS Parc National de Hwange (Zimbabwe) comme site d’étude. Le parc de Hwange est un environnement caractérisé par un fort risque de prédation pour les zèbres, qui font face à une forte densité de lions 8.
Dans le cadre de la bourse de terrain SFE², je souhaitais développer une approche comparative, l’idée étant d’étudier une population de zèbres sans prédateurs de manière similaire à ce que je vais réaliser sur la zone de Main Camp à Hwange (figure 1). Le parc national des Matopos, à environ 350 km au sud de Hwange (figure 1) offrait cette opportunité mais la végétation trop dense en cette période de l’année ne m’a pas permis d’obtenir les données que j’espérais collecter (photo 1). Par conséquent, j’ai choisi de réaliser des prospections dans d’autres zones du pays : le parc national de Kazuma Pan, le parc national de Zambezi et la zone ouest du parc national de Hwange (figure 1). Aucun zèbre n’a été vu dans le parc national de Kazuma Pan et seulement 3 groupes ont été vu dans le parc national de Zambezi, dont 2 pour lesquels la végétation trop dense n’a pas permis de déterminer les tailles de groupes et le recrutement avec exactitude (photo 2). Pour des raisons de visibilité, j’ai donc finalement choisi de me concentrer uniquement sur la zone ouest du parc national de Hwange qui est la zone qui offre la meilleure visibilité pour observer les zèbres.
-
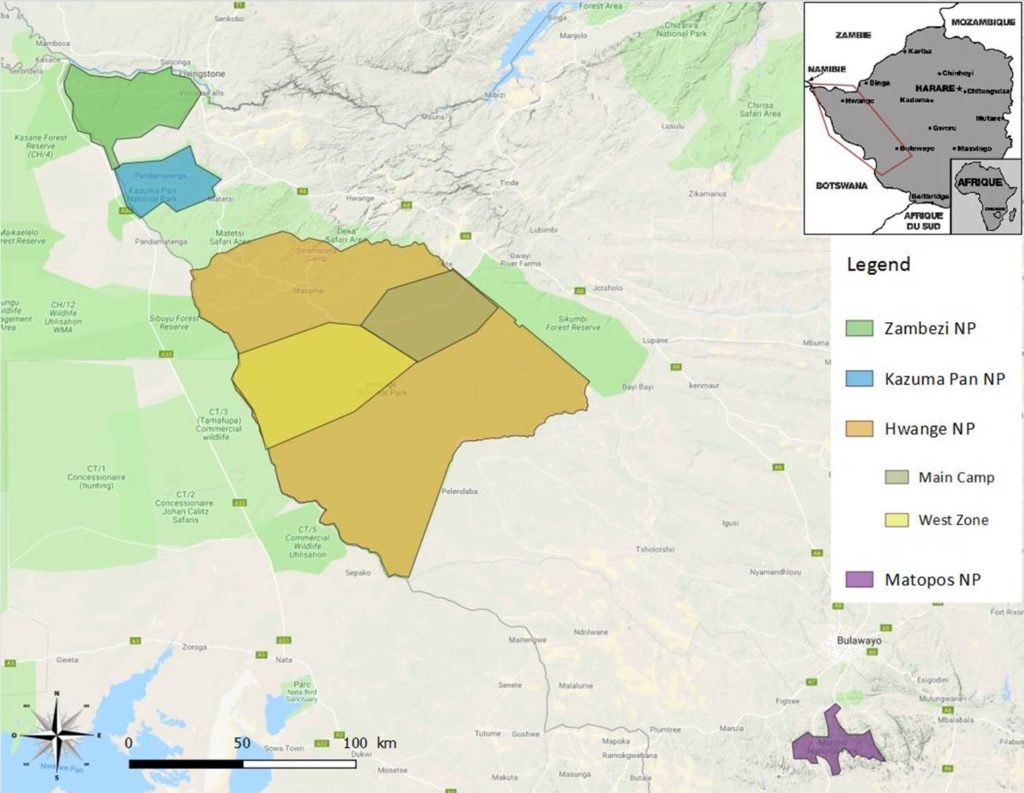
- Fig. 1. Carte montrant le site d’étude principal (Main Camp) et les autres sites qui ont pu être prospectés grâce à la bourse de terrain SFE² (c) Camille Vitet
-

- Photo 1. Végétation du parc national de Matopos (c) Camille Vitet

-

- Photo 2. Groupe de zèbres observé au parc national de Zambezi dans une végétation trop dense pour que la taille de groupe et le recrutement soient déterminés avec précision (c) Camille Vitet

Les groupes de la zone ouest sont migrateurs puisque, en saison sèche, l’absence d’eau dans cette zone les obligent à migrer vers la zone de Main Camp où des points d’eaux sont pompés et donc constamment approvisionnés en eau tout au long de l’année. En saison des pluies, ces groupes migrent vers la zone ouest, où le risque de prédation est plus faible que dans la zone de Main Camp1. Il est possible que cette migration soit une stratégie comportementale permettant de réduire le risque de prédation (« predation risk avoidance hypothesis ») 9,10. Cette hypothèse prédit un taux de survie des individus migrateurs, particulièrement des juvéniles, plus élevé que celui des individus résidents. Pour les groupes migrateurs de la zone ouest, on s’attend donc à (1) des tailles de groupe plus petites, et (2) d’avantage de poulains comparés aux groupes résidents de la zone de Main Camp, puisqu’ils font face à un taux de prédation plus faible lorsqu’ils migrent vers la zone ouest du parc en saison des pluies (période principale de naissance des poulains). En comparant les tailles de groupe et le recrutement des zèbres de la zone ouest avec ceux de la zone de Main Camp, l’étude s’oriente vers une question de dilution par effet de migration, la zone ouest constituant une zone refuge potentielle contre les prédateurs.
Méthodes
Pour chaque groupe de zèbres rencontré, la taille du groupe a été notée ainsi que le nombre de poulains au sein du groupe, qui informe sur le taux de recrutement. Un jeune est considéré comme « poulain » lorsqu’il est âgé de moins d’un an, ce qui se traduit par une taille au garrot d’au moins 10 cm plus courte que celle de sa mère 11. Quand cela était possible, des photos du côté droit et gauche de chaque individu ont également été prises afin de pouvoir les identifier l’année suivante à partir du motif de leurs rayures et ainsi évaluer leur taux de survie. Les photos ont également permis d’identifier les groupes déjà vu et ainsi d’éliminer les doublons dans les données.
Résultats et discussion
La taille d’échantillon pour les groupes de la zone ouest étant très faible (n=4), aucune analyse robuste ne peut être réalisée. Néanmoins, les résultats obtenus suggèrent que les tailles de groupes ne sont pas différentes entres les groupes migrateurs de la zone ouest et les groupes résidents de la zone de Main Camp (figure 2).
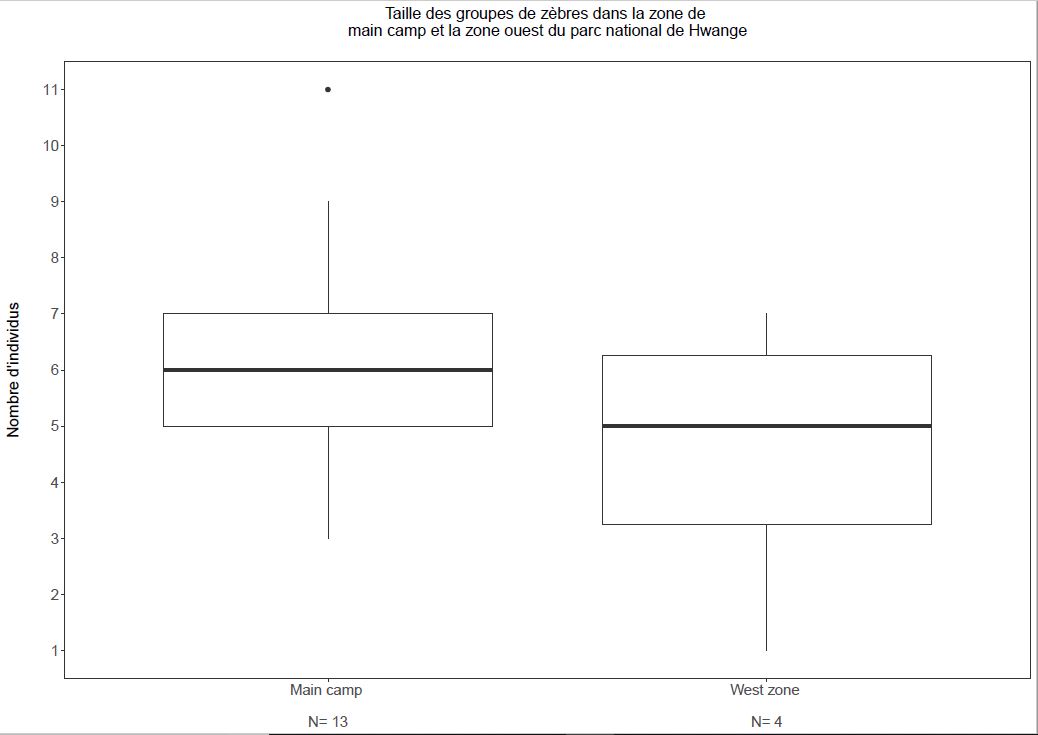
Fig. 2. Taille des groupes de zèbres de la zone de Main Camp et de la zone ouest
Ce résultat va à l’encontre de notre hypothèse et des résultats obtenus par Hebblewhite & Merrill chez le wapiti Cervus canadensis qui ont montré des tailles de groupes 20% plus large chez les groupes résidents comparés aux groupes migrateurs12. De plus, contrairement à ce qui était attendu, il semble que le taux de recrutement soit plus faible pour les zèbres migrateurs de la zone ouest (figure 3). Ce résultat contredit ceux d’une étude menée chez l’élan Alces alces qui a montré un taux de survie juvénile 2,6 à 2,9 fois plus élevé pour des individus migrateurs comparés à des individus résidents 13, mais appuie les résultats de l’étude menée par Hebblewhite & Merrill chez le wapiti où la survie des juvéniles étaient plus faible pour les individus migrateurs par rapport aux individus résidents12.
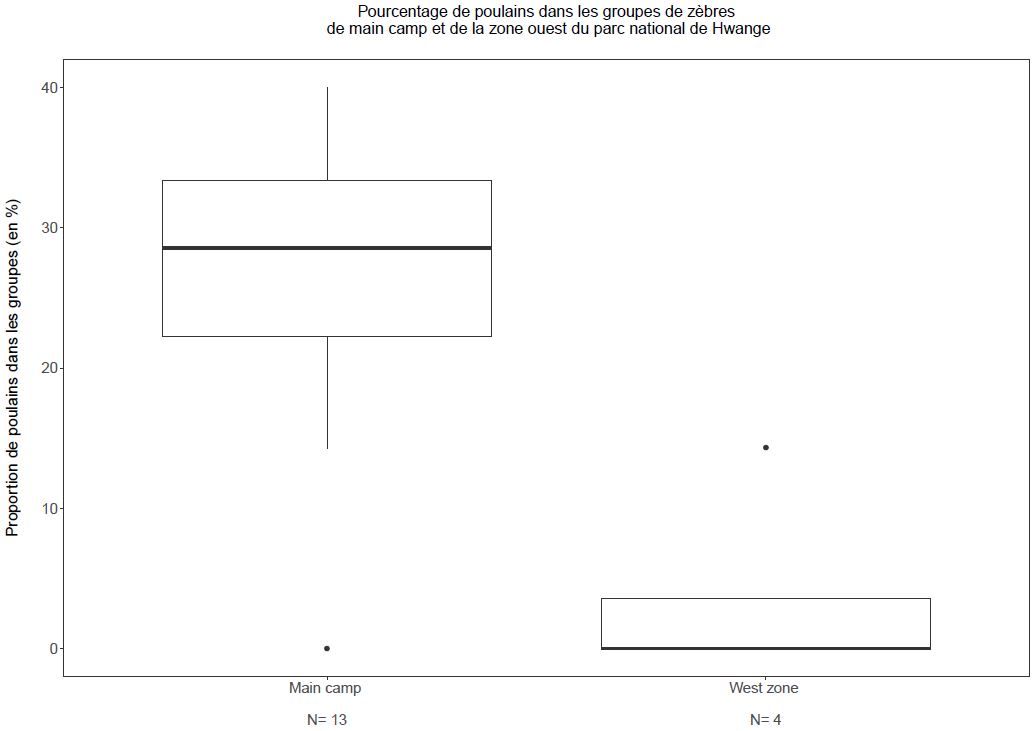
Fig. 3. Pourcentage de poulains au sein des groupes de zèbres de la zone de Main Camp et de la zone ouest
D’après ces résultats, il semble que le coût lié à la migration dans la zone ouest soit plus important que le bénéfice qu’elle devrait apporter en termes de réduction du risque de prédation. Plusieurs explications sont possibles pour expliquer ce taux de recrutement plus faible chez les groupes migrateurs: le coût énergétique du trajet peut affaiblir les individus, les ressources de la zone ouest peuvent être moins abondantes/riches, ou il est possible que le risque de prédation augmente lors de la migration, comme cela a été montré chez le cerf de Virginie Odocoileus virginianus, le cerf élaphe Cervus elaphus, et le cerf hémione Odocoileus hemionus14-16. Également, les groupes résidents de la zone de Main Camp pourraient utiliser les zones proches des habitations humaines (inexistantes dans la zone ouest) comme refuge contre les prédateurs à plus fine échelle (« human shield hypothesis »)17 ce qui expliquerait leur taux de recrutement plus élevé. En effet, des études ont déjà montrés que des stratégies comportementales à fine-échelle telles que celle d’utiliser les zones proches d’habitations humaines, peuvent diminuer le risque de prédation d’autant, voire plus, que des migrations à grande échelle18-20.
Davantage de données de taille de groupes et de recrutement sont nécessaires pour les groupes de la zone ouest afin de réaliser une analyse plus robuste. Cette analyse permettra de mieux comprendre l’impact du risque de prédation sur la démographie de cette espèce et de confirmer ou non les coûts qui semblent liés à la migration dans la zone ouest. Si les hypothèses sont validées (c.-à-d. les tailles de groupes sont plus petites et le recrutement est plus élevé pour les groupes migrateurs de la zone ouest), les résultats appuieront la « predation risk avoidance hypothesis » qui suggère un bénéfice de la migration en termes de réduction du risque de prédation. En revanche, si les résultats déjà obtenu se confirment (c.-à-d. les tailles de groupes sont similaires entre les deux zones et le recrutement est plus faible pour les groupes migrateurs de la zone ouest), se posera alors la question de savoir pourquoi certains groupes de zèbres migrent dans la zone ouest malgré le coût qui semble lié à la migration. Une hypothèse possible est que la zone ouest offre de meilleures ressources pour les zèbres, ce qui encourageraient certains groupes à migrer dans cette zone (« forage maturation hypothesis »)10,21.
Références
1. Loveridge, A. J., Searle, A. W., Murindagomo, F. & Macdonald, D. W. The impact of sport-hunting on the population dynamics of an African lion population in a protected area. Biol. Conserv. 134, 548–558 (2007).
2. Caro, T. M. (Timothy M. . Antipredator defenses in birds and mammals. (University of Chicago Press, 2005).
3. Lehtonen, J. & Jaatinen, K. Safety in numbers: the dilution effect and other drivers of group life in the face of danger. Behav. Ecol. Sociobiol. 70, 449–458 (2016).
4. Beauchamp, G. Disentangling the various mechanisms that account for the decline in vigilance with group size. Behav. Processes 136, 59–63 (2017).
5. Cresswell, W. & Quinn, J. L. Predicting the optimal prey group size from predator hunting behaviour. J. Anim. Ecol. 80, 310–319 (2011).
6. Markham, A. C., Gesquiere, L. R., Alberts, S. C. & Altmann, J. Optimal group size in a highly social mammal. Proc. Natl. Acad. Sci. 112, 14882–14887 (2015).
7. Majolo, B., de Bortoli Vizioli, A. & Schino, G. Costs and benefits of group living in primates: group size effects on behaviour and demography. Anim. Behav. 76, 1235–1247 (2008).
8. Grange, S. et al. Demography of plains zebras (Equus quagga) under heavy predation. Popul. Ecol. 57, 201–214 (2015).
9. Bergerud, A. T., Ferguson, R. & Butler, H. E. Spring migration and dispersion of woodland caribou at calving. Anim. Behav. 39, 360–368 (1990).
10. Fryxell, J. M. & Sinclair, A. R. E. Causes and consequences of migration by large herbivores. Trends Ecol. Evol. 3, 237–241 (1988).
11. Penzhorn, B. L. Age determination in Cape Mountain Zebras (Equus Zebra Zebra) in the Mountain Zebra National Park. Koedoe 25, 89–102 (1982).
12. Hebblewhite, M. & Merrill, E. H. Demographic balancing of migrant and resident elk in a partially migratory population through forage-predation tradeoffs. Oikos 120, 1860–1870 (2011).
13. White, K. S., Barten, N. L., Crouse, S. & Crouse, J. Benefits of migration in relation to nutritional condition and predation risk in a partially migratory moose population. Ecology 95, 225–237 (2014).
14. Nelson, M. E. & Mech, L. D. Wolf predation risk associated with white-tailed deer movements. Can. J. Zool. 69, 2696–2699 (1991).
15. Hebblewhite, M. & Merrill, E. H. Multiscale wolf predation risk for elk: Does migration reduce risk? Oecologia 152, 377–387 (2007).
16. Nicholson, M. C., Bowyer, R. T. & Kie, J. G. Habitat Selection and Survival of Mule Deer: Tradeoffs Associated with Migration. J. Mammal. 78, 483–504 (1997).
17. Berger, J. Fear, human shields and the redistribution of prey and predators in protected areas. Biol. Lett. 3, 620–623 (2007).
18. Anderson, A. P. et al. Summer resource selection scale-dependent by reintroduced elk in Wisconsin, USA. J. Wildl. Manage. 69, 298–310 (2005).
19. Johnson, C. J., Parker, K. L., Heard, D. C. & Gillingham, M. P. A Multiscale Behavioral Approach to Understanding the Movements of Woodland Caribou. Ecol. Appl. 12, 1840–1860 (2002).
20. Johnson, C. J., Parker, K. L., Heard, D. C. & Gillingham, M. P. Movement parameters of ungulates and scale-specific responses to the environment. J. Anim. Ecol. 71, 225–235 (2002).
21. Hebblewhite, M., Merrill, E. & McDermid, G. A Multi-Scale Test Of The Forage Maturation Hypothesis In A Partially Migratory Ungulate Population. Ecol. Monogr. 78, 141–166 (2008).
Commentaires récents