Limites nutritionnelles, stratégies d’acquisition et succès reproductif des plantes dans les pelouses
André Jardim Arruda
Email: ajarruda@gmail.com
Résumé
La production de graines est une étape cruciale du cycle de vie des spermatophytes (plantes avec graines), reflétant le potentiel de dispersion, de persistance de l’espèce et de la dynamique de la végétation. Les connaissances sur les filtres environnementaux liés au succès de la reproduction, y compris la viabilité et la qualité des graines, sont d’une extrême importance pour soutenir les stratégies de conservation, ainsi que les interventions de réhabilitation des zones dégradées. La disponibilité des nutriments dans le sol est l’un des principaux filtres environnementaux lié au succès de la reproduction et de l’établissement initial des plantes, et qui en conséquence influence la distribution des plantes et la dynamique de la succession végétale. Également, la mycorhization, ou l’association symbiotique racine/champignon, qui permet une meilleure absorption de l’eau et des nutriments du sol, est un autre facteur crucial pour le succès de la reproduction de plantes. Parmi les éléments que les mycorhizes aident à absorber, il se distingue le phosphore (P), un élément chimique peu mobile dans le sol et qui est crucial pour la production initiale de radicules après la germination et par conséquent pour le succès de l’établissement initial des plantes. Les études de la qualité des graines sont ainsi très importantes dans le domaine de la restauration écologique. La présente étude a testé des hypothèses liées à la qualité des graines pour cinq espèces végétales sur des parcelles aux sols avec différents niveaux de limitation nutritionnelle, en particulier du phosphore. La région de La Crau, localisée dans le sud de la France, est une pelouse semi-naturelle riche en espèces et est associée à des sols naturellement pauvres en éléments nutritifs. Dans un tel environnement, le P est un facteur limitant clé et les plantes ont développé des stratégies remarquables pour l’acquisition efficace de P par des associations avec des champignons mycorhiziens. De faibles teneurs en P dans le sol peuvent limiter fortement l’allocation des ressources aux graines, ce qui peut expliquer la faible capacité de régénération naturelle observée pour certaines espèces végétales dans ces milieux. L’objectif de ce travail était d’étudier l ‘influence de la fertilité des sols et de l’intensité des associations mycorhiziennes sur la qualité des graines dans les communautés végétales. Notre hypothèse est que le contenu nutritionnel du sol et les différentes stratégies d’acquisition du P par des plantes ont une relation directe avec la qualité et le contenu nutritionnel des graines. La région de La Crau abrite une végétation herbacée de grande richesse (localement nommée Coussoul). Dans la région, on peut distinguer des pelouses secondaires (friches) sur des parcelles anciennement cultivées en maraichage ou céréales entre les années 1980 et 2000 et abandonnées à des périodes différentes, ce qui se reflète directement dans les disponibilités contrastées de P. Les pratiques culturales (amendements et labour) ont modifié directement et durablement les caractéristiques physico-chimiques du sol. Pour cette étude nous avons choisi de comparer deux types de zones: Friche (trois parcelles) ou Coussoul (préservé). Nos résultats à ce jour montrent que les sols du Coussoul sont légèrement plus acides et contiennent beaucoup moins phosphore disponible pour les plantes que les friches. Nous avons aussi constaté une tendance à des pourcentages plus élevés de mycorhization des plantes du Coussoul. L’apport en phosphore modifie donc indirectement certains paramètres écophysiologiques de la végétation en diminuant la mycorhization, ce qui peut également être une conséquence de la dégradation du sol et des microorganismes du sol dans les friches de la Crau. Nous n’avons pas pu mettre en évidence un effet parcelle et donc un effet de la quantité de phosphore dans le sol sur la viabilité des graines pour les cinq espèces étudiées. Apparemment, la faible disponibilité de phosphore dans le sol de Coussoul (0.07 mg/g) peut être potentiellement compensée par le développement de relations entre des mycorhizes et les racines des plantes; ou bien cela se résume à d’autres conséquences écologiques, telles que la réduction de la quantité de fruits produite par individu. Les résultats des analyses nutritionnelles des graines (en cours) pourront fournir des informations complémentaires sur la qualité des graines, comme par exemple le contenu de phosphore. De plus, des taux élevés de graines non viables semble être une caractéristique marquante dans les écosystèmes associés aux sols sévèrement appauvris en P mais que dominés par des espèces pérennes, et donc un problème moins prononcé sur des écosystèmes dominés par des espèces annuelles telles que dans le Coussoul de La Crau.
Introduction
La production de graines est une étape cruciale du cycle de vie des plantes spermatophytes, reflétant le potentiel de dispersion, de persistance de l’espèce et de changement de la végétation. Les graines sont des éléments clés pour comprendre la dynamique de la régénération et de l’évolution des écosystèmes (Standish & Hobbs 2010; Hopper et al., 2016). De nombreuses communautés végétales sont limitées en graines, ce qui signifie que les microsites où les graines peuvent arriver et germer restent vacantes, ce qui peut être lié à une production de graines limitée ou à une faible qualité de graines (Clark et al. 1998). Les connaissances sur les filtres environnementaux liés au succès de la reproduction, y compris la viabilité et la qualité des graines, sont d’une extrême importance pour soutenir les stratégies de conservation, ainsi que les interventions pour la réhabilitation des zones dégradées. La disponibilité des nutriments dans le sol est l’un des principaux filtres environnementaux liés au succès de la reproduction et de l’établissement initial des plantes, et qui en conséquence influencent la distribution des plantes et la dynamique de la succession végétale. Ainsi, une meilleure compréhension de l’influence de la limitation du P dans le sol et des stratégies d’acquisition des nutriments dans la production des graines pourrait fournir des informations cruciales pour soutenir les pratiques de restauration et conservation de la biodiversité (Standish & Hobbs 2010; Hopper et al., 2016). Également, la mycorhization, association symbiotique racine/champignon, qui permet une meilleure absorption de l’eau et des nutriments du sol, est un autre facteur crucial pour le succès de la reproduction de plantes. Parmi les éléments que les mycorhizes aident à absorber, se distingue le phosphore (P): un élément chimique peu mobile dans le sol et qui est crucial pour la production initiale de radicules après la germination et par conséquent pour le succès de l’établissement initial des plantes. La présente étude vise à tester des hypothèses liées à la qualité de graines pour cinq espèces de plantes sur des parcelles aux sols avec différents niveaux de limitation nutritionnelle, en particulier du phosphore. L’objectif de ce travail était d’étudier l’influence de la fertilité des sols et de l’intensité des associations mycorhiziennes sur la qualité des graines dans les communautés végétales de la région de La Crau, qui abrite une végétation herbacée de grande richesse, étant donc importante pour la conservation de la biodiversité méditerranéenne. Notre hypothèse est que le contenu nutritionnel du sol et les différentes stratégies d’acquisition du P par des plantes ont une relation directe avec la qualité et le contenu nutritionnel des graines.
Méthodes
Zone d’étude
La Crau est une plaine plus de 45.000 ha située dans le sud de la France qui abrite une végétation herbacée de grande richesse (Buisson et Dutoit 2006). Le climat dans la région est méso-méditerranéen, caractérisé par trois mois d’été secs et chauds, des hivers pluvieux avec une température moyenne de 7°C et une température moyenne annuelle de 14,5°C. La pluviométrie annuelle moyenne varie entre 400 et 600mm (Buisson et al., 2004). Les parcelles choisies pour la présente étude sont situées dans la Réserve des Coussouls de Crau (Photo 1), qui est principalement constituée de pelouses sèches ou pseudo-steppes associée à des sols peu profonds et naturellement pauvres en éléments nutritifs, surtout en P. Dans la réserve, on peut distinguer la végétation naturelle (Coussoul) associée à plusieurs zones géologiques correspondant à différentes périodes de formation des sols, et des pelouses secondaires sur des parcelles anciennement cultivées en maraichage ou céréales entre les années 1980 et 2000 et abandonnées à des périodes différentes (Friches ; Photo 2), ce qui se reflète directement dans les disponibilités contrastées en P . Les pratiques culturales ont modifié directement et durablement les caractéristiques physico-chimiques du sol: e.g. phosphore et potassium sont toujours présents en plus grande quantité dans les friches, même dizaines d’années après abandon. Pour cette étude nous avons choisi de comparer deux types de zones: Friche et Coussoul.
-

- Photo 1. Réserve des Coussouls de Crau, qui est principalement constituée de pelouses sèches ou pseudo-steppes associée à des sols peu profonds et naturellement pauvres en éléments nutritifs, surtout en phosphore (P).
-

- Photo 2. Certaines friches ont été cultivées en céréales et/ou maraichage (melon) entre les années 1980 et 2000. Les pratiques culturales ont modifié directement et durablement les caractéristiques physico-chimiques du sol: e.g. phosphore et potassium sont toujours présents en plus grande quantité dans les friches, même dizaines d’années après abandon.
Pour la zone de friches, trois parcelles ont été sélectionnées et sont nommées selon les bergeries les plus proches: Couliès, Peau de Meau (PDM) et Valigne (Val). Le Coussoul préservé situé à proximité de chaque friche a également été échantillonné et les échantillons ont été rassemblés. Au final, nous comparons quatre parcelles: le Coussoul préservé et trois friches. Au total, cinq espèces des plantes ont été étudiées: Avena barbata, Senecio vulgaris, Hedypnois cretica, Geranium molle et Dactylis glomerata. Dans la suite du rapport, afin de l’alléger, elles seront nommées par leur nom de genre (car il n’y a qu’une espèce par genre).
Collection du sol, de racines et de graines
Les analyses nutritionnelles de sol ont été réalisées à partir de trois échantillons de sol pour chaque parcelle. Les échantillons ont été prélevés à 0-10 cm de profondeur dans chaque parcelle et tamisés (<2 mm) pour éliminer les éléments grossiers, les racines et les débris organiques. La colonisation mycorhizienne a été évaluée à partir des racines recueillies au minimum de trois individus de chaque espèce par parcelle. Les collectes de graines ont été exécutées entre Avril et Juillet 2018. Toutes les graines ont été récoltées avec des gants, pour éviter la contamination pour les futures analyses nutritionnelles.
Analyse de laboratoire
Les échantillons de sols ont été analysés par le Laboratoire Teyssier en évaluant les paramètres suivants: pH, P total, P disponible, carbone et azote. Le P assimilable dans les échantillons de sol a été mesuré selon la méthode décrite par Olsen et al. (1954). La colonisation par AMF a été évaluée à partir des racines collectées. Les racines après séchage ont été colorées en utilisant la méthode de Vierheilig et al. (1998). Pour le pourcentage total de la colonisation des racines par AMF, les hyphes internes, les vésicules et les arbuscules ont été comptés en utilisant la méthode des intersections étendues en utilisant 100 intersections (McGonigle et al., 1990). Les graines collectées ont été amenées au laboratoire et séchées dans un four à des températures entre 60°C et 70°C pendant 24 h (Atkinson, 1973). Les graines séchées ont été stockées dans un endroit sec, aéré et à l’abri de la lumière. Nous avons pesé avec une balance de précision quatre répliques × 25 graines par parcelle et par espèce soit au total 400 graines par espèce. Puis ces mêmes graines ont servi pour les tests de viabilité des graines (Photo 3). Nous avons ensuite pesé minimum 0.2 g de graines pour quatre répliques par parcelles nécessaire pour les futures analyses nutritionnelles. Pour l’analyse des graines, deux paramètres ont été évalués: la qualité de l’embryon à travers le test tetrazolium et le contenu nutritionnel à travers la méthode de spectrométrie d’émission optique couplée au plasma (ICP-OES) (Rodushkin et al., 1999). Le test tétrazolium est une méthode pour estimer la viabilité de l’embryon (Marcos-Filho 2015). La méthodologie ICP-OES permet l’analyse simultanée d’une large gamme d’oligo-éléments, pour lesquels nous attendons toujours les résultats qui son en train d’être collectés à University of Western Australia au mois de janvier/février 2019. Pour cela, quatre répétitions de 25 graines choisies au hasard parmi graines récoltées sur un minimum de 10 individus, totalisant l’analyse de 100 graines par espèce cible. Pour l’analyse du contenu nutritionnel des graines, la teneur en phosphore par rapport au poids sec de la graine sera évaluée, étant celui-ci un bon indice d’investissement de la graine en carbone (Grubb et Coomes, 1997), en plus d’autres d’oligo-éléments.
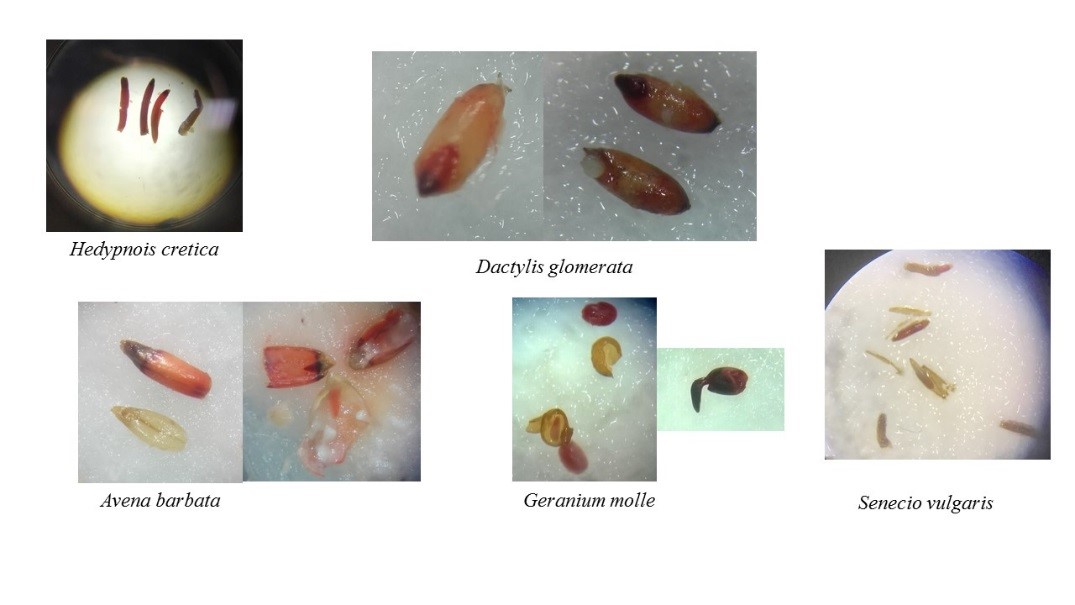
Photo 3. Photos de graines des 5 espèces observées à la loupe binoculaire après le test au tétrazolium.
Résultats et discussion
Les analyses de sol ont montré que la quantité de phosphore dans les sols des friches est environ 3 à 5 fois supérieure à celle du Coussoul (Figure 1 et Tableau 1). Aucune différence significative n’a été trouvée en ce qui concerne les teneurs en carbone, azote et pH. On a également trouvé une tendance à la diminution des associations mycorhiziennes pour les plantes poussant dans les friches (Figure 2). L’apport en phosphore, même ancien, peut donc modifier indirectement certains paramètres écophysiologiques de la végétation en diminuant la mycorhization, ce qui peut également être une conséquence de la dégradation du sol et des microorganismes dans les friches. Les graines d’Avena, Geranium et Senecio n’ont pas un poids significativement différent dans les différentes zones (Figure 3). Le poids des graines de Dactylis et Hedypnois est significativement plus faible sur la friche de Couliès. Pour Hedypnois le poids est significativement plus élevé sur la friche de Peau de Meau. Tant dis que pour Dactylis, le poids est significativement plus élevé sur la friche de Valigne. Cependant, il n’a pas un lien clair qui montre une tendance entre la quantité de phosphore dans le sol et le poids des graines lorsqu’on considère les cinq espèces. En ce qui concerne la viabilité des graines, Dactylis a significativement la plus faible, avec seulement 34.1% des graines viables (Figure 4). Les viabilités des graines de Geranium et Hedypnois sont significativement les plus élevées des cinq espèces (respectivement 88.3% et 81.0%). Avena et Senecio ont des viabilités intermédiaires (respectivement 67.6% et 71.3%). Aucune des cinq espèces étudiées n’a présenté de différence significative de viabilité en fonction des zones (friches ou coussouls) par rapport à la quantité de phosphore dans le sol.
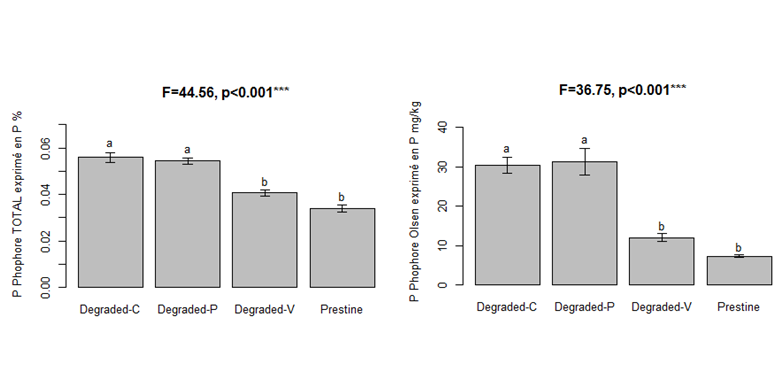
Figure 1: Phosphore total et disponible pour les plantes dans les sols du Coussoul (Pristine) des friches (Degraded-C, P and V)
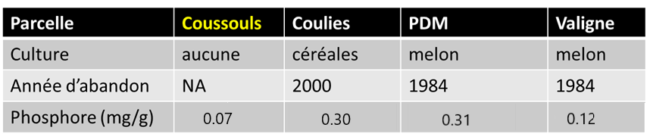
Tableau 1: Historique de dégradation et disponibilité de Phosphore du sol La Crau
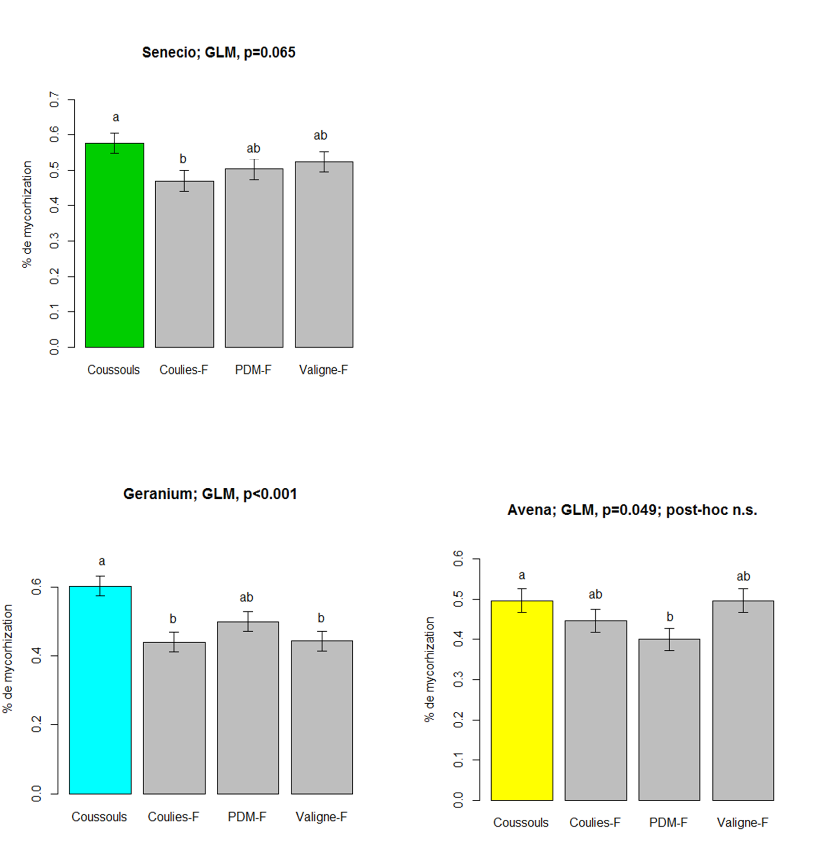
Figure 2: Taux de mycorhization pour trois espèces dans le Coussouls et les friches (Coulies-F, PDM-F et Valigne-F).
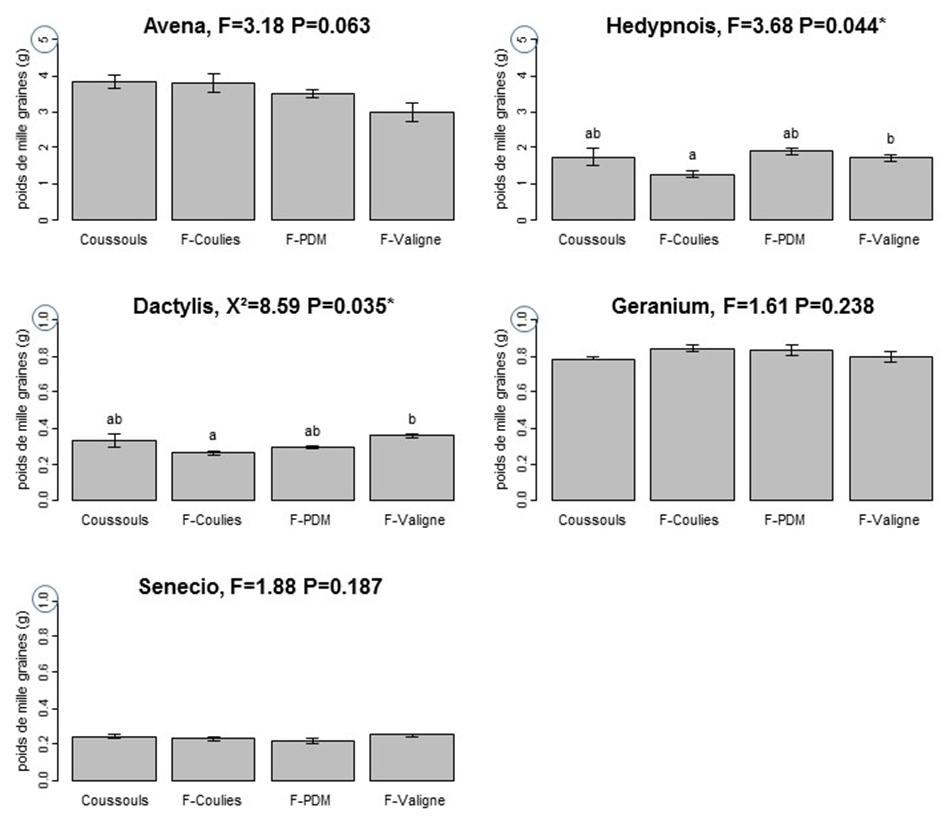
Figure 3: Représentation graphique du poids de mille graines (g) en fonction des zones étudiées, pour 5 espèces. Les axes des ordonnées sont compris entre 0 et 5g pour Avena et Hedypnois et entre 0 et 1 g pour Dactylis, Senecio et Geranium. Pour Dactylis, nous avons effectué un test non paramétrique Kruskal-Wallis suivi d’un test post-hoc pairwise de Wilcoxon avec ajustement de Bonferroni. Pour les quatre autres espèces, nous avons effectué le test ANOVA suivie d’un test post-hoc de Tukey.

Figure 4: Pourcentage de viabilité des graines en fonction des cinq espèces étudiées. Nous avons effectué le test ANOVA suivie d’un test post-hoc de Tukey
Apparemment, la faible disponibilité du phosphore dans le sol de Coussoul (0.07 mg/g) peut être potentiellement compensée par le développement de relations mycorhiziennes; ou bien cela se résume à d’autres conséquences écologiques, telles que la réduction de la quantité de graines produites par individu. Les résultats de l’analyses nutritionnelles qui seront bientôt obtenus peuvent fournir des informations complémentaires sur la qualité des graines. Les connaissances sur les filtres environnementaux liés aux processus de régénération naturelle de ces paysages et qui servent à accroître les connaissances sur l’évolution de ces écosystèmes, sont d’une extrême importance pour soutenir les stratégies de conservation, ainsi que les interventions pour la réhabilitation des zones dégradées. Les études de la qualité des graines sont un point très important pour l’écologie et en particulier pour la restauration écologique qui nécessite d’avoir des espèces avec des graines de bonnes qualités afin de réussir au mieux la restauration de la zone concernée.
Références bibliographiques
Atkinson D (1973) Some general effects of phosphorus deficiency on growth and development. New phytologist 72:101-111 Buisson E, Dutoit T (2004) Colonisation by native species of abandoned farmland adjacent to a remnant patch of Mediterranean steppe. Plant Ecology 174:371-384 Buisson E, Dutoit T (2006) Creation of the Natural Reserve of La Crau: implications for the creation and management of protected areas. Journal of Environmental Management 80:318-326 Clark JS, Macklin E, Wood L (1998) Stages and spatial scales of recruitment limitation in southern Appalachian forests. Ecological Monographs 68:213–235 Grubb PJ, Coomes DA (1997) Seed mass and nutrient content in nutrient-starved tropical rainforest in Venezuela. Seed science research 7:269-280 Hopper SD, Silveira FAO, Fiedler PL (2016) Biodiversity hotspots and Ocbil theory. Plant Soil 403:167-216 Marcos-Filho J. (2015) Seed vigor testing: an overview of the past, present and future perspective. Scientia Agricola 72:63-374. McGonigle TP, Miller MH, Evans DG, Fairchild GL, Swan JA (1990) A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytologist 115:495-501 Olsen S, Cole C, Watanabe F, Dean L (1954) Estimation of available phosphorus in soils by extraction with sodium bicarbonate. USDA Circular Nr 939, US Gov. Print. Office, Washington, D.C. Rodushkin, I., Ruth, T., Huhtasaari, A (1999) Comparison of two digestion methods for the elemental determination in plant material by ICP techniques. Anal. Chim. Acta 378, 191–200. Standish R, Hobbs R (2010) Restoration of OCBILs in southwestern Australia: response to Hopper. Plant soil, 330:15–18
Commentaires récents