La Société Française d’Ecologie (SFE) vous propose cette semaine le regard de Frédéric Médail, Alex Baumel, Katia Diadema et Jérémy Migliore, chercheurs à l’Institut Méditerranéen de Biodiversité et d’Ecologie (IMBE) et au Conservatoire Botanique de Porquerolles, sur les écosystèmes méditerranéens.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions après cet article. Les auteurs vous répondront et une synthèse des contributions sera ajoutée après chaque article.
La biodiversité végétale méditerranéenne,
organisation et évolution
Frédéric Médail1, Alex Baumel1, Katia Diadema2 et Jérémy Migliore1
1 : Institut Méditerranéen de Biodiversité et d’Ecologie (IMBE, UMR-CNRS 7263), Aix-Marseille Université. frederic.medail@imbe.fr
2 : Conservatoire botanique national méditerranéen de Porquerolles, Hyères, France
Regard R38, édité par Anne Teyssèdre
( Fichier PDF )
Mots clés : biogéographie, changements globaux, climat, endémisme, évolution, paléoécologie, phylogéographie, plantes,
région méditerranéenne, spéciation, zones refuges
———
L’exceptionnelle richesse biologique des écosystèmes méditerranéens a été soulignée depuis une quinzaine d’années, grâce à la démarche mondiale basée sur le concept de point-chaud de biodiversité* (hotspot) terrestre. Tel est le cas des plantes à fleurs et fougères, puisqu’environ 10 % des végétaux supérieurs du globe se rencontrent en région méditerranéenne, sur une surface seulement égale à 1,6 % de la superficie terrestre (Médail & Quézel, 1997). Estimée à 25000 espèces ou 30000 espèces et sous-espèces, la richesse floristique se concentre en deux principaux pôles, l’un occidental qui comprend la péninsule ibérique et le Maroc, et l’autre oriental avec la Turquie et la Grèce. Les îles et les montagnes déterminent des secteurs biogéographiques originaux, où le taux d’endémisme dépasse souvent 20 %. Sur la base de la richesse et de l’endémisme végétal, il a été possible de définir dix points-chauds régionaux de biodiversité méditerranéenne, abritant environ 5500 végétaux endémiques (44 % de l’ensemble) sur 22 % des terres, soit environ 515 000 km2 (Médail & Myers 2004, cf. Figure 1).

Figure 1 : Points-chauds (hotspots) régionaux de biodiversité végétale de la région méditerranéenne
(d’après Médail & Quézel 1997, complété)
A une échelle locale (inférieure ou égale à 0,1 ha), les écosystèmes méditerranéens se caractérisent par des richesses floristiques très hétérogènes, selon les types d’habitats considérés, mais certains comme les pelouses à espèces annuelles sont parmi les plus riches communautés connues. Pour tenter d’expliquer cette forte biodiversité, l’étude de l’influence des processus écologiques locaux a été initialement privilégiée. Les stress environnementaux et les perturbations ont en effet un rôle majeur dans la dynamique des systèmes biologiques méditerranéens, en maintenant de fortes hétérogénéités spatiales et temporelles. Mais la mosaïque des paysages, les fortes variations topographiques et enfin l’impact très ancien de l’homme sur les milieux naturels modèlent aussi cette biodiversité (Blondel et al., 2010). Plus récemment, une meilleure prise en compte des paramètres biogéographiques* et évolutifs montre toute l’importance des processus macroécologiques*, qui s’inscrivent dans de vastes échelles spatiales et temporelles.
Glossaire
Biogéographie : discipline ayant pour objet l’étude de la répartition des êtres vivants à la surface du globe et la compréhension des processus qui régissent cette répartition.
Endémisme : phénomène caractérisant la distribution restreinte à une zone biogéographique donnée d’une espèce, d’un genre ou d’un groupe systématique plus vaste (famille, ordre..).
Gondwana : super-continent issu de la fragmentation de la Pangée vers 200 Ma et regroupant l’Afrique, l’Arabie, l’Inde, l’Amérique du Sud, l’Antarctique et l’Australie.
Horloge moléculaire : théorie selon laquelle une séquence génétique évolue à une vitesse à peu près constante au cours du temps, la distance séparant deux séquences permettant de dater l’époque de divergence des êtres vivants auxquels elles appartiennent. Différentes techniques analytiques permettent de corriger l’hétérogénéité des taux d’évolution moléculaire entre lignées génétiques, afin d’avoir une échelle temporelle de l’évolution.
Hotspot : cf. Point chaud de biodiversité
Laurasie : super-continent issu de la fragmentation de la Pangée vers 200 Ma et regroupant l’Amérique du Nord, le Groenland, l’Europe centrale et du Nord et l’Asie.
Macroécologie : discipline qui analyse les facteurs et processus écologiques se déroulant à de vastes échelles spatiales (depuis la région biogéographique jusqu’au système Terre), et qui étudie les grands équilibres et cycles de la biosphère.
Paléoécologie : discipline visant à estimer l’organisation et la dynamique des espèces ou communautés vivantes passées, parallèlement à la quantification de certaines variables environnementales (paramètres climatiques, régimes de feu, pH des eaux lacustres, etc.), grâce à la localisation, l’identification, la quantification et la datation de restes biologiques (assemblages fossiles, grains de pollen, charbons et cernes de bois, foraminifères, etc.).
Paléogéographie : étude de la géographie des continents au cours des différentes époques géologiques, elle s’appuie sur la théorie de la tectonique des plaques qui a mis en évidence le perpétuel changement de la configuration des masses continentales.
Point-chaud de biodiversité (biodiversity hotspot) : secteur géographique de concentration exceptionnelle en espèces végétales ou animales et en espèces endémiques.
Phylogénétique : analyse des relations de parentés entre différents êtres vivants, en vue de comprendre l’évolution des organismes. Actuellement, les arbres phylogénétiques sont le plus souvent reconstitués à partir des données moléculaires.
Phylogéographie : étude des principes et phénomènes gouvernant la distribution géographique de diverses lignées génétiques, en particulier au sein d’une même espèce ou entre espèces proches parentes. La phylogéographie est appliquée à différentes échelles spatiales, et elle intègre la biogéographie, la phylogénétique mais aussi la paléoécologie, afin d’examiner la généalogie du monde vivant selon un cadre spatio-temporel précis.
Refuge : voir Zone refuge.
Spéciation : processus évolutif d’apparition de nouvelles espèces. On distingue plusieurs types de spéciation, basés sur la répartition géographique des populations en divergence au cours du processus (les spéciations allopatrique, péripatrique, parapatrique et sympatrique).
Taxonomie : discipline de classification des organismes vivants, qui a pour objet de les décrire et de les regrouper en entités appelées taxons, afin de les identifier, les nommer et enfin les classer, en s’appuyant sur les principes de la systématique (science organisant le classement des taxons et leurs relations).
Zone refuge : aire géographique particulière (souvent des îles, montagnes ou gorges) où des lignées génétiques distinctes ont persisté au fil des fluctuations climatiques passées (cycles glaciaires par exemple), du fait de caractéristiques environnementales favorables.
La diversité végétale méditerranéenne, fruit d’une histoire contrastée
La flore méditerranéenne actuelle est formée d’un mélange complexe d’espèces aux origines biogéographiques* variées et plus ou moins anciennes. Elle est le résultat de différenciations locales à partir d’espèces ancestrales, et de multiples migrations de végétaux, répétées au fil du temps. L’histoire géologique particulièrement mouvementée de cette région et les fortes variations climatiques survenues depuis 2 millions d’années (Ma) constituent des facteurs historiques clés pour expliquer cette biodiversité très hétérogène (Thompson, 2005).
Située à la charnière entre les vestiges des deux anciens « super-continents », la Laurasie* et le Gondwana*, séparés depuis 200 Ma, la région méditerranéenne a été peuplée à partir de ces deux ensembles. Son fond floristique se compose d’éléments de souche méridionale (tropicale), principalement d’affinité asiatique, et d’éléments extra-tropicaux, de souche autochtone ou septentrionale. L’individualisation d’îles et bras de terre liée au déplacement de diverses microplaques tectoniques dans cette région depuis au moins 140 Ma (début du Crétacé) a rendu possible de nombreux échanges entre ces deux ensembles. Cette paléogéographie* complexe a également permis le développement d’une riche flore endémique, à la faveur de l’isolement plus ou moins marqué des différentes régions, et de l’extrême hétérogénéité des substrats, de la géomorphologie, mais aussi du climat (Verlaque et al., 1997).
L’origine de la flore méditerranéenne remonte au Néogène, qui est la deuxième partie de l’Ere Tertiaire ou Cénozoïque. C’est en effet au Miocène (entre 23 et 5 Ma) et surtout au Pliocène (entre 5 et 1,8 Ma), que se diversifient bon nombre de végétaux structurants (pins, chênes, olivier, filaires, cistes, etc.) qui imposent la réalité d’une végétation, au moins localisée, de type méditerranéen. Ces taxons étaient alors associés à des végétaux très exigeants en température (espèces dites mégathermes), aujourd’hui cantonnés aux régions tropicales.
Entre 3,5 et 2,4 Ma se met en place le climat méditerranéen qui se caractérise par un renforcement du contraste des saisons et un abaissement des températures hivernales. Au début du Pléistocène, il y a environ 1,8 Ma, des variations drastiques de température ont joué un rôle majeur dans l’extinction des végétaux tempérés-chauds qui prospéraient jusqu’alors. Toutefois, certains arbres relictuels plus mésophiles (i.e. adaptés à des températures plus modérées), tels que le Cotonéaster de l’Atlas, le Genévrier thurifère et l’If, persistent encore aujourd’hui, en particulier dans plusieurs zones refuges* tertiaires du pourtour méditerranéen. Finalement, les pulsations climatiques du Quaternaire (alternance des cycles glaciaires-interglaciaires) ont grandement influencé les dynamiques des paysages, les migrations ou les isolements successifs des flores entre les diverses régions, en permettant l’émergence de nouvelles espèces.
Les analyses biogéographiques* de végétaux méditerranéens à vaste distribution (Laurier, Olivier, Bruyère arborescente, Myrte) témoignent du rôle de carrefour biogéographique de la région méditerranéenne, et de la diversité des processus en œuvre (voir encadré ci-dessous).
L’histoire biogéographique* du Myrte commun en Méditerranée
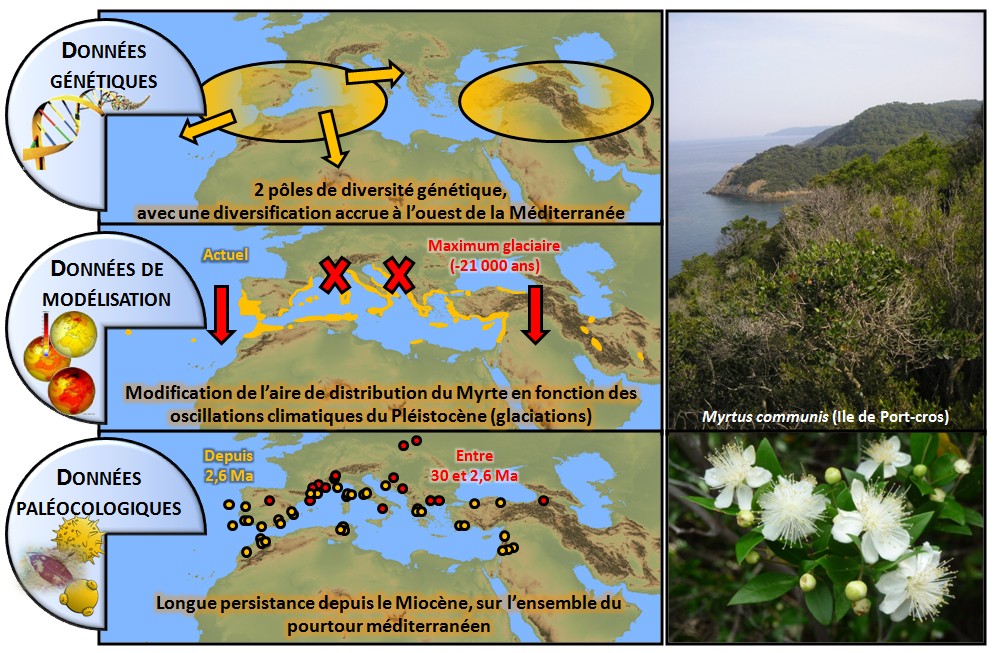
Si l’on veut bien comprendre l’origine et la mise en place de la flore méditerranéenne à l’échelle de l’ensemble de la région, il est nécessaire d’étudier des végétaux à vaste distribution et dont les données fossiles sont disponibles, car elles permettent d’estimer les tempos évolutifs (via l’horloge moléculaire*). Une telle approche a été récemment conduite pour étudier la distribution géographique des lignées génétiques du Myrte commun (Myrtus communis, Myrtacée), buisson caractéristique des maquis thermophiles méditerranéens (Migliore et al. 2012, cf. Figure 2 ci-dessous).

Figure 2 : Résultats synthétiques issus des approches de paléobotanique, génétique et modélisation bioclimatique
conduites sur le modèle Myrte commun (Myrtacée) (d’après Migliore et al., 2012).
Les archives paléobotaniques (fossiles, pollens) révèlent l’origine ancienne du Myrte, présent au moins depuis le milieu du Tertiaire (Oligocène), soit au minimum 30 Ma, et sa distribution sur tout le pourtour méditerranéen au cours du Pléistocène (entre 1,8 Ma et 12.000 ans BP). L’étude de la distribution et des liens de parenté entre séquences moléculaires par la phylogéographie* met en évidence deux événements majeurs de diversification du Myrte : l’un à la transition Miocène-Pliocène (vers 5-6 Ma), période caractérisée par la « crise de salinité du Messinien » consécutive à la fermeture du détroit de Gibraltar ; l’autre, au Pléistocène, avec la mise en place progressive du climat méditerranéen et des cycles de glaciation. On note aussi la divergence engendrée par l’isolement des populations est-méditerranéennes et la plus grande diversification des populations occidentales.
La modélisation de la niche écologique basée sur les données de distribution actuelle du Myrte, et projetée avec les données climatiques reconstituées du dernier maximum glaciaire (il y a 21 000 ans), souligne l’impact des glaciations dans l’extinction des populations les plus nordiques et le glissement de l’aire de distribution de l’espèce vers les latitudes plus basses, aux conditions environnementales plus favorables.
La place clé des zones refuges du pourtour méditerranéen
Depuis la formalisation par J. Haffer, à la fin des années 1960, de la théorie générale des zones refuges* basée sur l’exemple amazonien, le rôle joué par ces zones dans la distribution de la biodiversité actuelle a été maintes fois démontré, tant à l’échelle globale qu’à l’échelle régionale (ex. Hewitt, 2004). En Méditerranée, la forte hétérogénéité environnementale a permis à bon nombre de végétaux de trouver un habitat favorable lors des multiples bouleversements climatiques passés.
Les études de paléoécologie* et de phylogéographie* confirment bien que les zones refuges* forment des territoires-clés pour expliquer l’organisation et l’originalité de la biodiversité méditerranéenne. Ces refuges abritent des espèces reliques et des lignées végétales anciennes, et ils se localisent notamment : (i) dans certaines régions méridionales épargnées par le front glaciaire, notamment les grandes péninsules (Ibérie, Italie, Balkans, Anatolie) et les îles, et (ii) dans les secteurs où la topographie (massifs isolés, gorges, falaises, vallons côtiers) et/ou le climat local particulier (importante humidité atmosphérique) ont facilité la persistance de ces flores anciennes ; un des exemples les plus remarquables est celui des vallons côtiers humides et chauds d’Andalousie méridionale qui abritent des populations végétales reliques d’origine tertiaire (ex. Rhododendron ponticum, Prunus lusitanica, Psilotum nudum…), vestiges des forêts laurifoliées du Tertiaire.

Figure 3 : Principales zones refuges de la région méditerranéenne mises en évidence par les études phylogéographiques
(d’après Médail & Diadema, 2009).
L’étude d’un jeu de données génétiques des populations méditerranéennes de plus de quatre-vingt espèces a permis l’identification et la délimitation d’une cinquantaine de zones refuges péri-méditerranéennes (Médail & Diadema, 2009) (Figure 3). Trente-trois sont situées dans la partie occidentale du bassin et dix-neuf à l’Est. Il existe une congruence spatiale significative entre ces cinquante-deux refuges et les zones majeures de biodiversité méditerranéenne : la totalité des territoires à fort taux d’endémisme* végétal est incluse dans les refuges* identifiés et la moitié des refuges est englobée dans les dix points-chauds régionaux de richesse floristique (Figures 1 et 3). Ces résultats suggèrent que des mécanismes évolutifs et biogéographiques* similaires ont eu une influence déterminante sur la biodiversité actuelle, et expliquent les congruences entre secteurs riches en endémiques et zones d’originalité génétique.
Les résultats phylogéographiques* accumulés depuis une dizaine d’années montrent aussi la complexité du fonctionnement de ces zones refuges, dont l’influence fluctue selon leur localisation vis-à-vis des fronts glaciaires et dépend des épisodes paléoenvironnementaux considérés. On peut même trouver de nettes disparités de réponse entre lignées génétiques, au sein d’une même espèce. Ainsi, chez le Hêtre, Magri et al. (2006) ont montré que les différents cycles glaciaires-interglaciaires n’avaient pas eu le même impact sur la persistance locale ou la colonisation des lignées, certaines recolonisant l’Europe sur des milliers de kilomètres après la dernière glaciation, alors que d’autres furent bloquées au bout des grandes péninsules, mais pas forcément par les montagnes ! Les implications évolutives de ces processus complexes sont également déterminantes, car en situation de refuge les populations de végétaux méditerranéens ont pu survivre durant plusieurs cycles de glaciations, migrer et se subdiviser durant les glaciaires, mais se réunir et s’hybrider pendant les interglaciaires.
L’endémisme* méditerranéen : « beaux vestiges » et « jeunes étoiles »
Les paléontologues nous apprennent que l’extinction des espèces est un phénomène évolutif courant, les espèces ou les genres ayant une persistance relativement « courte », de quelques millions d’années. L’un des moyens pour résister aux changements environnementaux ou aux catastrophes est d’avoir une vaste distribution géographique.
Comment expliquer alors le taux d’endémisme* élevé de la flore méditerranéenne ? Deux hypothèses sont envisageables, celle de taux d’extinction plus faibles (hypothèse des refuges*) et celle de taux de diversification plus forts (hypothèse des points-chauds de spéciation*).
Selon la première hypothèse, des plantes endémiques anciennes (paléoendémiques), souvent reliques d’aires autrefois plus vastes, auraient survécu dans des zones d’une grande stabilité climatique depuis le Tertiaire. Cette première hypothèse s’appuie donc sur des phénomènes de stabilité écologique mais aussi démographique grâce à des adaptations facilitant la persistance (régénération végétative, clonalité, bulbes ou rhizomes, dispersion à courte distance).
La deuxième hypothèse s’appuie sur le modèle de Chown (1997) selon lequel la probabilité de spéciation* croît avec la diminution de l’abondance et avec la fragmentation de l’habitat ; ainsi, la rareté et l’endémisme* seraient à la fois causes et conséquences de la spéciation*. D’origine récente, les néoendémiques vivraient sur le lieu de leur origine. Tel est le cas des cyclamens endémiques, apparus pour la plupart il y a moins de trois millions d’années. D’autres taxons méditerranéens très typiques, tels que les cistes, montrent également une diversification relativement récente, comparée aux taxons reliques du Tertiaire.

Figure 4 : Habitat de la Sabline de Provence, Arenaria provincialis (Caryophyllacée), endémique de Basse-Provence (© F. Médail)
Diverses études phylogénétiques* récentes permettent bien de vérifier que la limite Pliocène-Pléistocène (vers 1,8 Ma), période d’aridification puis de refroidissement, a favorisé la spéciation* en région méditerranéenne. C’est le cas de la Sabline de Provence (Arenaria provincialis), petite Caryophyllacée localisée aux collines de la région marseillaise, qui est, du point de vue de son histoire, une « néoendémique » (Youssef et al., 2011). Son origine remonterait au début du Pléistocène, à partir d’un ancêtre d’affinité alpine affectionnant des climats plus froids que l’actuel. La forte diversité génétique de ses populations, sur un territoire si exigu, semble appuyer l’hypothèse d’une grande stabilité démographique. La Sabline vit actuellement dans les lapiazs et les éboulis, milieux rocheux méditerranéens, où la stabilité écologique est assurée par une succession végétale très lente, une grande hétérogénéité locale de la topologie offrant de multiples refuges* dans le voisinage (Figure 4).
Actuellement, de nombreuses plantes endémiques des zones côtières et des petits massifs proches du littoral font face à un changement nouveau de leur environnement, l’urbanisation, dont l’intensité est bien supérieure à celle des changements climatiques passés et actuels. L’étroite concordance géographique des pressions humaines (zones densément peuplées) et des zones refuges abritant souvent de nombreuses plantes endémiques à distribution restreinte, conduit à une situation inédite de crise écologique pour cette biodiversité unique. Pour le seul département des Alpes-Maritimes, près de la moitié des zones refuges* se situe là où les pressions d’urbanisation sont les plus élevées (Médail & Diadema, 2006)…
Vers une prise en compte multi-facette de la biodiversité méditerranéenne,
pour mieux la conserver
Mieux comprendre les phénomènes environnementaux qui agissent sur les espèces constitue une étape importante pour conserver la biodiversité. En région méditerranéenne, où les conséquences biologiques des changements globaux seront probablement très contrastées (Blondel & Médail, 2009), il s’agit en particulier d’évaluer les processus démographiques et fonctionnels des populations végétales et animales. Cependant, la forte imprévisibilité des événements climatiques et des autres perturbations qui caractérisent cette région complique les simulations démographiques au sein de ces écosystèmes hétérogènes et changeants (Blondel et al., 2010).
Même s’il a traversé avec plus ou moins de succès le crible de grandes crises, cet héritage biologique s’avère fragilisé par la rapidité et l’ampleur des changements environnementaux en cours et prévus par les diverses modélisations. L’ensemble du bassin méditerranéen ne devrait pas être affecté de manière homogène sur le plan climatique (Hoerling et al., 2012), et l’étude du passé nous montre que bon nombre d’espèces ont pu trouver un habitat favorable lors des anciennes variations de climat. En premier lieu, l’identification précise de la localisation des zones refuges* méditerranéennes constitue une priorité en conservation biogéographique*, car ces zones abritent, on l’a vu, une biodiversité unique, sur le plan taxonomique (espèces endémiques), évolutif (forte diversité et originalité génétiques) et fonctionnel (émergence de nombreuses adaptations locales). Dans une perspective de conservation évolutive de la biodiversité (Nieto-Feliner, 2011), il convient aussi de bien comprendre les processus de réponse des végétaux (extinction, migration ou persistance) face aux changements globaux du passé, afin de mieux estimer ceux du futur.
S’il n’est bien évidemment pas envisageable de contrôler les conditions climatiques, l’étude des diverses influences liées aux perturbations ou aux changements du mode d’usage des terres pourrait également nous apporter des informations complémentaires et permettre de définir des propositions de gestion plus efficaces.
Les politiques de conservation en région méditerranéenne devraient donc considérer deux objectifs complémentaires : (i) la protection des espaces (habitats remarquables), qui permettrait la préservation de zones uniques au monde (par exemple les centres d’endémisme*) incluant des espèces endémiques et/ou relictuelles et des zones refuges*, sans oublier les corridors d’habitats garants des connectivités biologiques ; (ii) la protection des espèces, indispensable pour la conservation des végétaux menacés dont la distribution est plus éparse et ne présente pas de tendance à l’agrégation. Cette double approche doit être mise en œuvre en priorité sur la frange littorale, la plus menacée (Blondel & Médail, 2009). Néanmoins, et contre toute attente, les zones littorales, même parmi les plus urbanisées (par exemple la Principauté de Monaco, les caps), recèlent parfois encore quelques « trésors » de biodiversité végétale, pour peu que persistent quelques fragments d’habitats semi-naturels.
Ainsi, seules des approches intégrées de conservation biogéographique* et évolutive, analysant conjointement les différentes gammes d’échelles spatio-temporelles et les multi-facettes de la biodiversité, depuis les paysages jusqu’aux gènes, permettront de fournir des critères et indicateurs utiles pour une conservation globale des écosystèmes et des ressources génétiques du bassin méditerranéen, en incluant les interactions avec les sociétés et les territoires
—-
Bibliographie
Blondel J. & Médail. F., 2009. Biodiversity and conservation. In : Woodward J.C. (ed.). The physical geography of the Mediterranean. Oxford University Press, Oxford, pp. 615-650.
Blondel J., Aronson J., Bodiou J.-Y. & Boeuf G., 2010. The Mediterranean region. Biological diversity in space and time, 2nd ed. Oxford University Press, Oxford.
Chown S.L., 1997. Speciation and rarity: separating cause from consequence. In : Kunin W.E. & Gaston K.J., (eds.). The biology of rarity. Chapman and Hall, London, pp. 91-109
Hewitt G.M., 2004. Genetic consequences of climatic oscillations in the Quaternary. Philosophical Transactions of the Royal Society of London B, 359 : 183-195.
Hoerling M., Eischeid J., PerlwitzJ., Quan X., Zhang T. & Pegion P., 2012. On the increased frequency of Mediterranean drought. Journal of Climate, 25 : 2146-2161.
Magri D., Vendramin G.G., Comps B., Dupanloup I., Geburek T., Gömöry D., Latałowa M., Litt T., Paule L., Roure J.M., Tantau I., Van Der Knaap W.O., Petit R.J. & Beaulieu de J.-L., 2006. A new scenario for the Quaternary history of European beech populations: palaeobotanical evidence and genetic consequences. New Phytologist, 171 : 199–221.
Médail F. & Diadema K., 2009. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. Journal of Biogeography, 36 : 1333-1345.
Médail F. & Myers N., 2004. Mediterranean Basin. In : Mittermeier R.A., Robles Gil P., Hoffmann M., Pilgrim J., Brooks T., Mittermeier C.G., Lamoreux J. & da Fonseca G.A.B. (eds.). Hotspots revisited: Earth’s biologically richest and most endangered terrestrial ecoregions. CEMEX (Monterrey), Conservation International (Washington) & Agrupación Sierra Madre (Mexico), pp. 144-147.
Médail F. & Quézel P., 1997. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Annals of the Missouri Botanical Garden, 84 : 112-127.
Migliore J., Baumel A., Juin M. & Médail F., 2012. From Mediterranean shores to central Saharan mountains: key phylogeographical insights from the genus Myrtus. Journal of Biogeography, 39 : 942-956.
Nieto Feliner G., 2011. Southern European glacial refugia: a tale of tales. Taxon, 60 : 365-372.
Thompson J.D., 2005. Plant evolution in the Mediterranean. Oxford University Press, Oxford.
Verlaque R., Médail F., Quézel P. & Babinot J.-F., 1997. Endémisme végétal et paléogéographie dans le bassin méditerranéen. Geobios, M.S. 21 : 159-166.
Youssef S., Baumel A., Véla E., Juin M., Dumas E., Affre L. & Tatoni, T., 2011. Factors underlying the narrow distribution of the Mediterranean annual plant Arenaria provincialis (Caryophyllaceae). Folia Geobotanica, 46 : 327-350.
—
Pour en savoir plus
Médail F., 2005. Mise en place et évolution de la biodiversité : l’exemple de la flore méditerranéenne. In : Marty P., Vivien F.D., Lepart J. & Larrère R. (eds.). Les biodiversités : objets, théories, pratiques. CNRS Editions, Paris, pp. 97-112.
Médail F. & Diadema K., 2006. Biodiversité végétale méditerranéenne et anthropisation : approches macro et micro-régionales. Annales de Géographie, numéro thématique « Les territoires de la biodiversité » 651: 618-649.
—–
Article édité par Anne Teyssèdre
Selon le modèle de Chown, la probabilité de spéciation croît avec la diminution de l’abondance et avec la fragmentation de l’habitat. Or les pressions d’urbanisation participent à la diminution de l’abondance et à la fragmentation de l’habitat. Est-il possible que les pressions d’urbanisation, si elles ne sont pas excessives, puissent être un facteur de l’accroissement du taux de spéciation sur le long terme?
Bonjour,
la plausibilité d’une différenciation croit avec l’isolement mais celle de l’extinction aussi, c’est donc difficile de d’émettre des hypothèses là-dessus, il faudrait modéliser le phénomène de diversification et d’extinction dans plusieurs milieux plus ou moins fragmentés et plus ou moins perturbés.
alex
Bonjour,
Je trouve ce regard rassurant : je m’imaginais en effet que l’urbanisation pléthorique et accélérée du Midi de la France et du littoral de l’Italie nuisait à la végétation au point de la détruire complètement (comme par exemple les merveilleux fonds sous-marins de mes jeunes années 1950 à 1960, qui ont été littéralement ratiboisés ). Quand je voyais les baous de Vence envahis par les villas, et la Costa Smeralda de Sardaigne transformée en un ridicule gazon verdoyant et bien arrosé mécaniquement, à l’usage de quelques personnes aux goûts douteux souvent dépourvues de tout sens de la beauté de la Nature, je désespérais. Je vois qu’il y a des zones refuges et que le myrte a un bel avenir, et cela me fait bien plaisir. Enfin, qui vivra verra, mais ce Regard n°38 me revigore un peu.
Quoique… Même si les espèces végétales rares et remarquables peuvent être préservées par les actions proposées dans cet article, l’invasion du maquis et des forêts méditerranéennes par les villas, routes et autres « infrastructures » m’inquiète toujours. Est-ce que la biologie de la conservation ne s’intéresse qu’aux espèces remarquables ou endémiques ?
Bonjour,
la biologie de la conservation s’intéresse à la diversité et surtout à préserver les processus sous jacents à cette diversité. La diversité à venir est selon moi imprévisible et peut émerger de « matériel » différents, populations d’espèces rares ou communes.
alex
En attendant la réaction des auteurs à ces commentaires et questions, j’aimerais répondre à la question finale de Marthe.
Non, bien sûr, les sciences de la conservation ne s’intéressent pas seulement aux espèces menacées, rares ou endémiques, ni aux seuls habitats remarquables, mais aussi à la biodiversité « ordinaire », c’est-à-dire aux populations, espèces et réseaux d’espèces communes.
Ceci, entre autres raisons, parce que l’apparition, la survie et l’évolution de toute espèce dépend du fonctionnement et de la résilience (capacité de retour à l’état antérieur après perturbation) du réseau écologique auquel elle appartient ; et ceci non pas seulement à l’échelle locale, mais à l’échelle plus vaste des paysages et écosystèmes régionaux, eux-mêmes en interaction à l’échelle des continents, des océans et de la biosphère dans son ensemble.
Les recherches en écologie des vingt dernières années montrent clairement que le fonctionnement et la résilience des écosystèmes dépend de la diversité des espèces qui les composent (voir à ce sujet le regard 3 sur cette plateforme). Et c’est bien sur cette diversité d’espèces, pour la plupart communes, que reposent les multiples fonctions et « services » fournis aux sociétés par les écosystèmes (cf. le regard 4). Le plaisir de contempler un beau paysage repose lui-même surtout sur la diversité de la nature « ordinaire », car les espèces rares ou menacées y sont par définition peu visibles.
Les recherches montrent aussi que l’intensité croissante des pressions (directes et indirectes) exercées par les humains sur de nombreux écosystèmes terrestres ou aquatiques réduit leur résilience et menace leur fonctionnement (voir par exemple les regards 30, 31 et 37).
De manière générale, les sciences de la conservation s’intéressent aux déterminants de la diversité biologique, à la structure et au fonctionnement des systèmes écologiques (populations, communautés, réseaux écologiques, écosystèmes..), ainsi qu’aux causes et conséquences de leurs changements rapides actuels, dans le but ultime de freiner ces changements. Etant donné que ces changements rapides sont massivement liés à l’expansion actuelle des sociétés humaines et de leurs activités sur l’ensemble de la planète, il s’agit donc de développer – et de faire connaître – des stratégies, outils et méthodes susceptibles de réduire l’impact de nos sociétés sur les écosystèmes et la biosphère, c.-à-d. en d’autres termes de « réconcilier » les humains et la nature (cf par ex. les regards 1, 4, 6, 7, 11, 13, 14, 19, 20, 21, 24, 27, 34, 36) [tout en prenant garde aux dérives et aux effets rebonds négatifs des stratégies incomplètes (cf. le regard 12)].
Livres conseillés :
– « Ecologie et biodiversité, des populations aux socioécosystèmes », D. Couvet et A. Teyssèdre, Belin 2010.
– « L’exigence de la réconciliation », sous la direction de C. Fleury et A-C. Prévot-Julliard, Fayard, 2012.
Bien cordialement,
Anne T.
Bonjour,
je voudrais savoir si dans le passé il y a eu des migrations floristiques de l’ouest du Bassin Méditerranéen (Espagne, Maroc) vers l’Est et vers l’Italie. Si oui, de quelles espèces et dans quel temps géologique ?
Y a-t-il des publications où on parle de ca?
Merci d’avance pour votre réponse.
Bonjour,
Si les migrations méditerranéennes ouest-est sont apparemment bien moins nombreuses que celles venant de Méditerranée orientale ou d’Asie, il y en a effectivement ; par exemple chez les Silene ou chez les Cistus :
Systematics, character evolution, and biogeography of Cistus L. (Cistaceae) based on ITS, trnL-trnF, and matK sequences par Beatriz Guzmán & Pablo Vargas, Molecular Phylogenetics and Evolution, 2005.
Mais les cas d’étude sont à développer pour mieux estimer la magnitude générale de ce type de migration !
F. Médail