La Société Française d’Ecologie (SFE) vous propose ce regard d’Evelyne Heyer, Professeur d’Anthropologie Génétique au Muséum National d’Histoire Naturelle, sur l’évolution humaine (évolution bioculturelle).
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
———-
Comment la culture agit sur notre évolution
par Evelyne Heyer,
Professeur d’Anthropologie Génétique au Muséum National d’Histoire Naturelle
Article édité par Anne Teyssèdre
——-
Mots clés : Evolution humaine, anthropologie, populations, culture, génétique, interactions bio-culturelles.
——–
- Introduction
- L’influence de la langue
- Organisation matrimoniale et structuration génétique des populations
- Transmission du succès de reproduction
- Un exemple d’évolution bioculturelle chez l’Homme
- Conclusion
- Glossaire
- Bibliographie
- Forum de discussion sur ce regard
——-
Introduction
L’espèce humaine est une. Les études génétiques ont montré sa forte homogénéité : en moyenne deux humains ne diffèrent que de un pour mille dans leurs génomes*, ou séquences d’ADN (c.-à-d. qu’un nucléotide* sur 1000 est différent, ce calcul n’incluant pas les insertions et délétions). A quoi sont dues ces différences ? Pas simplement aux pressions de l’environnement physique, mais aussi à l’environnement culturel et social. Ainsi la langue, les règles du mariage ou de la parenté, le statut social ou l’héritage économique, créent aussi des frontières symboliques entre les groupes, contribuant ainsi à façonner le profil génétique des individus. Autrement dit : la culture agit aussi sur l’évolution comme série d’études récentes l’ont mis en évidence.
Le premier aspect de cette influence concerne les règles ou coutumes de mariages. Dans de nombreuses sociétés, les mariages ont longtemps été contrôlés par des critères culturels : ils ont généralement lieu entre membres d’un même groupe social ou culturel caractérisé par le statut social (voire la caste) ou l’appartenance religieuse, comme chez les juifs, les musulmans et les chrétiens par exemple. Ou encore, la préférence pour les mariages entre cousins (mariages dits endogamiques, ou endogames) conduit à limiter la variabilité génétique des populations.
L’influence de la langue
Au delà des traditions, la langue est un critère culturel d’appariement entre les sexes, qui influe sur l’évolution et la différenciation génétique des populations. Elle tend à créer une plus grande proximité génétique pour cette raison simple : les mariages sont plus nombreux entre personnes partageant la même langue. Deux populations qui se mélangent par les mariages vont peu à peu se ressembler. Inversement, les populations qui échangent peu entre elles vont progressivement se différencier génétiquement. En Asie Centrale, nous avons pu mesurer précisément cette concordance entre proximité linguistique et génétique.
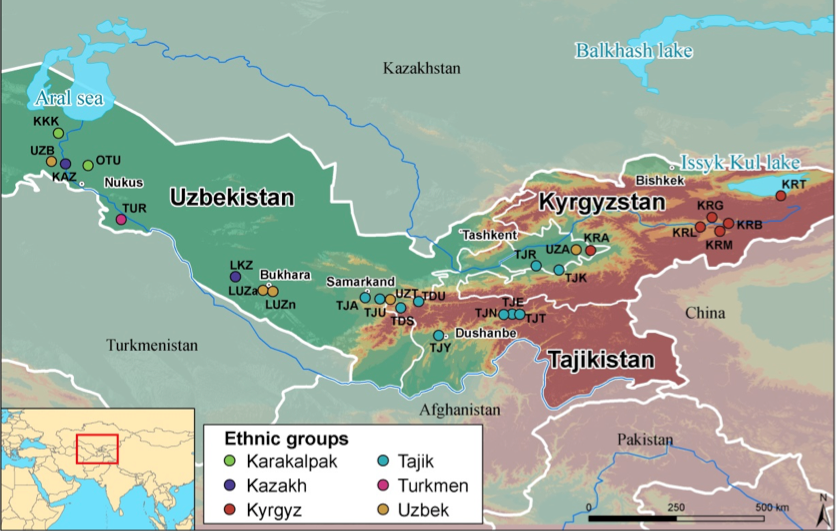
Fig.1 : Carte des 26 différentes populations échantillonnées. (Source : Heyer, Segurel et Hegay, 2014)
Dans cette région du monde, qui recouvre les cinq anciennes républiques de l’URSS : Turkmenistan, Ouzbekistan, Kirghizstan, Tajikistan et Kazakhstan, deux groupes de populations se côtoient : celles de langue Turk et celles de langue indo-iranienne. Dans chacune de ces populations, nous avons collecté des données à la fois linguistiques et génétiques. Les résultats montrent l’existence de différences génétiques entre les deux groupes, différence qui s’expliquent par la limitation des mariages entre ces groupes. Résultat plus intéressant : cette corrélation entre différences linguistiques et génétiques est indépendante de la géographie. Autrement dit, on préfère se marier avec quelqu’un proche linguistiquement, même s’il est éloigné géographiquement. Conclusion : ici, la langue renforce les différences génétiques entre les populations de langues différentes et augmente la proximité génétique entre populations de langue proche (Heyer and Mennecier, 2009).
L’influence désormais démontrée de la culture sur les variables génétiques des populations renverse ainsi le paradigme classique qui postule que ce sont les différences « biologiques » qui influent sur les différences culturelles entre groupes humains. Il reste que ces différences sont faibles. Ainsi, à l’échelle de l’Eurasie, les deux groupes d’Asie Centrale sont très proches génétiquement, ce qui montre bien que des échanges ont toujours eu lieu entre les populations malgré les barrières linguistiques (Martinez-Cruz, 2011).
Organisation matrimoniale et structuration génétique des populations
D’autres facteurs culturels que la langue contribuent à agir sur la variété génétique des populations : l’organisation sociale en est un. Dans le monde, la majorité des populations (70%) sont dites « patrilocales ». Autrement dit après le mariage le couple s’installe dans le village de l’époux et l’épouse migre. Ces migrations des femmes entre villages, sur un grand nombre de générations, peuvent se mesurer par la faible différenciation de l’ADN mitochondrial (1) entre villages.
Inversement, dans les populations matrilocales, où ce sont les hommes qui migrent dans le village de leur épouse, les différences génétiques entre les populations sont plus fortes sur le chromosome Y (masculin, et donc transmis uniquement de père en fils). On retrouve donc ici, l’impact d’un facteur culturel, la patri- ou matri-localité, sur la diversité génétique des populations (Seielstad et al., 1998; Hamilton et al., 2005).
En Asie Centrale, où les populations sont toutes patrilocales, on retrouve cet impact sur la diversité génétique : il y a moins de différences entre populations pour l’ADN mitochondrial (marqueur de lignée maternelle) que pour le chromosome Y (marqueur de lignée paternelle) (Chaix et al., 2007; Segurel et al., 2008).
(1) : Présents dans quasi toutes les cellules corporelles (dites somatiques) et dans les cellules reproductrices femelles (ovules) qui les transmettent à la génération suivante, mais absents des spermatozoïdes, les organites cellulaires que sont les mitochondries (sièges des réactions énergétiques liées à la respiration) et leur ADN sont des marqueurs de la lignée féminine. (Note de l’éditrice.)
Cette signature génétique des règles du mariage nous a par ailleurs permis d’estimer que les populations pygmées d’Afrique Centrale seraient régies par des règles matrilocales, et ce depuis un assez grand nombre de générations (Verdu et al 2013).
Après la langue et la règle de mariage (patri ou matri-locale), un autre facteur agit sur la variabilité génétique des populations : les règles de descendance. Ainsi, en Asie Centrale, les populations de langue Turk sont dites patrilinéaires, c’est–à-dire que la transmission du nom, de l’héritage, passe par la lignée par la voie paternelle. Par contre, les populations de langue indo-iranienne sont dites cognatiques, c’est-à-dire que chez elles la transmission du nom ou/et de l’héritage à la descendance est autant maternelle que paternelle. L’organisation patrilinéaire crée des lignées paternelles remontant à un ancêtre paternel éloigné de 7 à 10 générations. Plusieurs de ces lignages se regroupent en clans à partir d’un ancêtre paternel commun encore plus éloigné. Et les clans se regroupent à leur tour en tribus qui revendiquent un même ancêtre commun très éloigné, souvent mythique. Cette organisation patrilinéaire en lignage, clans, tribus, se retrouve dans les traces génétiques : on peut les observer sur le chromosome Y. Cette correspondance est intéressante : elle montre que les généalogies décrites oralement correspondent bien à des généalogies biologiques et atteste d’une organisation patrilinéaire qui existe depuis au moins 20-30 générations (Chaix et al, 2004).
La filiation patrilinéaire a des conséquences génétiques : elle réduit la diversité génétique au sein d’un groupe et augmente les différences entre les groupes. La figure 2 ci-dessous, qui distingue les populations selon leur mode de subsistance (agriculteurs sédentaires vs éleveurs nomades) et donc leur organisation sociale (patrilinéaire vs cognatique, voir le tableau 1), illustre ces résultats. Voilà donc un autre paramètre culturel qui agit sur la diversité génétique des populations.
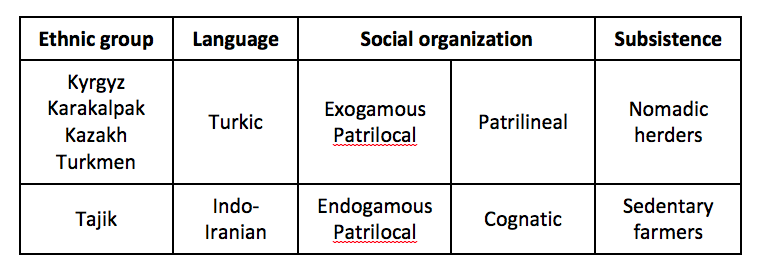
Tableau 1: Description schématique des différents groupes ethniques d’Asie centrale.
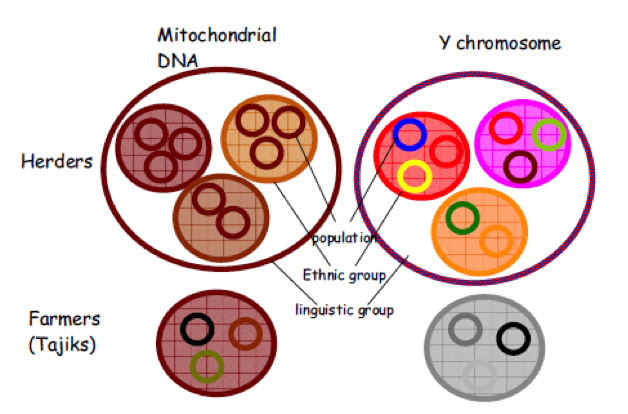
Fig.2 : Représentation schématique de la structuration génétique au niveau du groupe ethnique. Les différentes couleurs correspondent à des différences génétiques, les nuances de la même couleur indiquant de faibles différences. (Source : Heyer et al., 2009)
Transmission du succès de reproduction
Un autre facteur culturel agit sur l’évolution biologique d’une population: le succès reproducteur des individus, c’est-à-dire le nombre de leurs enfants qui atteignent l’âge de se reproduire à leur tour, et la variation de ce « succès reproducteur » entre individus. /Ce succès/ dépend notamment de facteurs culturels tels que la richesse qu’elle soit matérielle ou de relation sociale (Borgerhoff-Mulder et al, 2009) : plus on est « riche », plus des conditions de vie sont aisées, plus les chances sont grandes pour les enfants de survivre, de se marier et donc de se reproduire à leur tour, etc. C’est ainsi que les traits génétiques de familles aisées ont plus de chance de se transmettre que d’autres. Ce ne sont donc pas des avantages biologiques au sens strict qui jouent ici, mais simplement les conditions de vie matérielle plus avantageuses. On a donc ici un mécanisme de « sélection culturelle » qui agit indépendamment de la sélection naturelle.
Dès les années 1970, l’existence d’une transmission culturelle du succès de reproduction à la descendance – influant donc sur l’évolution génétique des populations – a été décrite par des anthropologues. Dans certaines populations amérindiennes d’Amazonie (Neel, 1970), les hommes de haut rang sont ceux qui ont le plus de femmes et de ce fait ont un plus grand nombre d’enfants. Ce fort succès reproducteur est transmis à la descendance : leurs fils sont également les plus polygames et ont un succès reproducteur plus élevé. Il y a donc bien transmission culturelle du succès reproducteur au fil des générations.
Si l’on se tourne vers certaines populations de Maoris de Nouvelle-Zélande, ce sont les femmes de haut statut social qui ont un plus fort succès reproducteur. En effet leurs enfants survivent mieux grâce à de meilleures conditions de vie. Le statut social est transmis aux filles qui ont pour les mêmes raisons, à leur tour, un plus fort succès reproducteur (Murray-McIntosh RP et al, 1998). Les données démographiques ont permis de repérer ce phénomène dans des populations rurales françaises du XVIIe siècle à nos jours : ici c’est la possession et l’héritage de terres qui favorise le succès reproducteur (Heyer, 1993). Dans les populations francophones de l’Est du Québec, le succès reproducteur est aussi transmis des parents aux enfants, en lien avec l’émigration (Heyer and Austerlitz, 2005). La « sélection culturelle » laisse donc des traces dans la diversité génétique des populations.
A l’aide d’une méthode génétique appelée théorie de la coalescence* (cf. figure 1 ci-dessous), qui permet de reconstituer la généalogie d’un échantillon de gènes en remontant le temps jusqu’à l’ancêtre commun de l’échantillon, nous avons mis au point un test permettant de détecter l’existence de cette transmission du succès reproducteur (Sibert et al, 2002, Brandenburg et al, 2012). L’application de ce test aux populations d’Asie Centrale a permis de mettre en évidence l’héritabilité du succès reproducteur dans les populations patrilinéaires (Heyer et al 2015, AJPA). Autrement dit, les hommes dotés d’un succès reproducteur élevé ont tendance à transmettre cette caractéristique -cet avantage sélectif- à leurs fils. Plus généralement, le succès reproducteur dépend d’un facteur culturel – la richesse ou le statut social – qui est transmis aux descendants.
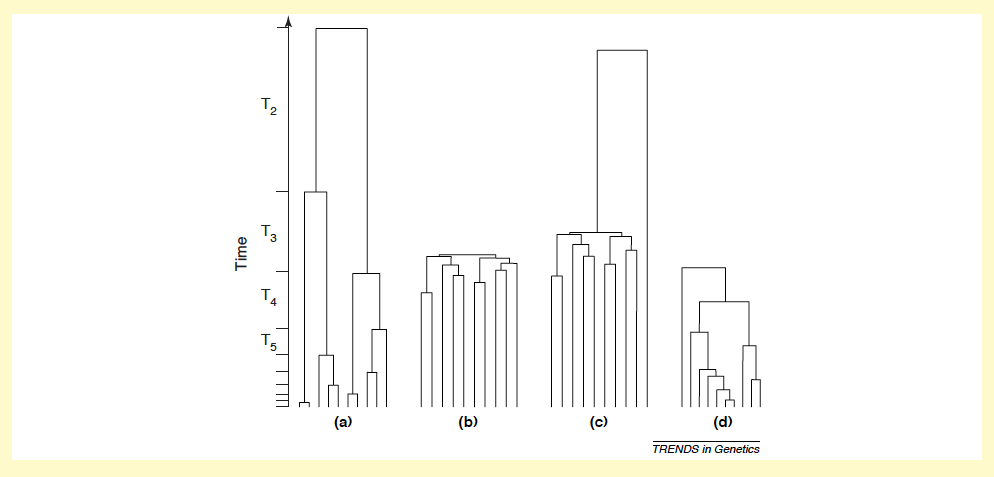
Fig.3 : Arbres de coalescence de gènes. (a) Généalogie d’un locus neutre dans une large population de taille constante.
(b) Locus neutre dans une population en expansion démographique. (d) Locus neutre dans une population avec transmission du succès reproducteur. Source : Heyer et al., 2005.
En résumé, plusieurs facteurs culturels contribuent à agir sur la génétique d’une population : la langue, les règles du mariage, le statut social, la transmission d’un rang ou d’une appartenance religieuse (pour une revue, voir Heyer et al. 2012). Ces phénomènes culturels ouvrent un nouveau pan sur le lien fort entre culture et évolution biologique chez l’Homme. Ils s’ajoutent à ce qui est classiquement appelé « évolution bio-culturelle » dans le cas où l’Homme modifie culturellement son environnement, auquel il s’adapte ensuite biologiquement (notion illustrée dans l’encadré ci-dessous).
Encadré: Un exemple d’évolution bio-culturelle chez l’Homme
Dans l’évolution biologique « simple », les individus sont sélectionnés par les pressions de l’environnement, par exemple parce qu’ils sont mieux adaptés génétiquement que d’autres à l’altitude, en région montagneuse. Dans certains cas, c’est l’Homme qui a modifié son environnement avec sa culture ; en retour ce nouvel environnement a agi sur lui en sélectionnant ceux qui étaient biologiquement les plus aptes à y vivre, lesquels ont ensuite transmis cette aptitude à leurs enfants.

Pasteur Afar, Erta Ale, Ethiopie.
© CC-BY-SA / Ji-Elle
C’est l’exemple classique de la tolérance au lactose : dans la prime enfance, le lait est notre nourriture de base. Mais, dès l’adolescence, 70 % des êtres humains le digèrent mal : la lactase, l’enzyme intestinale qui transforme le sucre du lait, ou lactose, en sucres utilisables, devient inopérante ou produite en trop faible quantité. Pourtant, certains, devenus adultes, continuent d’apprécier une forte consommation de lait. Dans certaines régions d’Afrique, de la péninsule arabique ou du Nord de l’Europe, 80 % des adultes tolèrent encore le lactose.
Des études ont montré que des modifications génétiques, des mutations, permettent de continuer à digérer le lait, l’enzyme lactase restant alors produite en suffisance. Il suffit que ces mutations deviennent avantageuses pour qu’elles se répandent dans la population au fil des générations. Or c’est le cas dans les groupes d’éleveurs dont l’alimentation est depuis plusieurs milliers d’années à base de lait frais, car les individus porteurs de ces mutations survivent et se reproduisent mieux que les autres. De fait, la tolérance au lactose est très fréquente dans ces populations traditionnellement pastorales.
De manière intéressante, les études génétiques ont aussi montré que ce sont des mutations différentes que l’on retrouve en Europe ou en Afrique. C’est un des exemples bien documentés d’évolution convergente (i.e., évolution de caractères biologiques semblables chez deux ou plusieurs lignées différentes, éventuellement très éloignées, sous la pression sélective de facteurs environnementaux (au sens large) similaires. Note de l’éditrice).
Conclusion
Partout et de tout temps, les groupes humains ont inventé, réinventé, modifié des traits culturels pour se différencier de leurs voisins. Ces traits culturels tendent à constituer des communautés plus ou moins homogènes, qui finissent par influer sur leur variété génétique,. La transmission culturelle ne signifie pas la fin de l’évolution biologique de l’espèce humaine ; elle contribue au contraire à agir sur elle et à la modifier.
Glossaire
- Génome : l’ensemble de l’ADN d’un individu, que ce soit la partie codante (les gènes) ou l’ADN non-codant. L’ADN d’un humain est formé de la succession de 3 milliards de nucléotides.
- Nucléotide : Molécule organique constituant l’élément de base d’un acide nucléique tel que l’ADN. Dans l’ADN, il peut prendre l’une des quatre formes – adénine, thymine, guanine et cytosine – souvent désignées comme les quatre « lettres » (A,T,G et C) de l’ADN.
- Théorie (et méthode) de la coalescence : En génétique, la théorie de la coalescence permet de reconstituer la généalogie d’un échantillon de séquences d’ADN homologues en remontant le temps jusqu’à l’ancêtre commun de l’échantillon. Deux séquences d’ADN sont d’autant plus proches dans l’arbre que leurs séquences se ressemblent.
Bibliographie
Borgerhoff Mulder M, Bowles S, Hertz T et al., 2009. Intergenerational wealth transmission and the dynamics of inequality in small-scale societies. Science, 326, 682–688.
Brandenburg JT, Austerlitz F, Toupance B., 2012. Impact of fertility transmission and other sociodemographic factors on reproductive success and coalescent trees. Genet Res (Camb). 2012 Jun;94(3):121-31
Chaix R, Austerlitz F, Khegay T. et al., 2004. The genetic or mythical ancestry of descent groups: lessons from the Y chromosome. American journal of human genetics 75(6):1113-1116.
Chaix R, Quintana-Murci L, Hegay T, Hammer MF, Mobasher Z, Austerlitz F, and E. Heyer, 2007. From social to genetic structures in central Asia. Current biology : CB 17(1):43-48.
Hamilton G, Stoneking M and L. Excoffier, 2005. Molecular analysis reveals tighter social regulation of immigration in patrilocal populations than in matrilocal populations. Proceedings of the National Academy of Sciences of the United States of America 102(21):7476-7480.
Heyer E., 1993. Population structure and immigration; a study of the Valserine Valley (French Jura) from the 17th century to today. Annals of Human Biology, vol 20, no 6, pp 565-573.
Heyer E, Sibert A & F. Austerlitz, 2005. Cultural transmission of fitness: genes take the fast lane. Trends in Genetics, 21, 234–239.
Heyer E & P. Mennecier, 2009. Genetic and linguistic diversity in Central Asia In: Hombert FdEaJ, editor. Becoming Eloquent. Amsterdam/Philadelphia: John Benjamins Publishing Company. p 163- 180
Heyer E, Chaix R, Pavard S, and F. Austerlitz, 2012. Sex-specific demographic behaviours that shape human genomic variation. Molecular ecology 21(3):597-612.
Heyer E, Brandenburg JT, M. Leonardi et al., 2015. Patrilineal populations show more male transmission of reproductive success than cognatic populations in Central Asia, which reduces their genetic diversity. American journal of physical anthropology. under print.
Martinez-Cruz B, Vitalis R, Segurel L. et al. . 2011. In the heartland of Eurasia: the multilocus genetic landscape of Central Asian populations. European journal of human genetics : EJHG 19(2):216-223.
Murray-McIntosh RP, Scrimshaw BJ, Hatfield PJ & D. Penny D, 1998. Testing migration patterns and estimating founding population size in Polynesia by using human mtDNA sequences. Proceedings of the National Academy of Sciences of the United States of America, 95, 9047–9052.
Neel JV, 1970. Lessons from a ‘‘primitive’’ people. Science, 170, 815–822.
Segurel L, Martinez-Cruz B, L. Quintana-Murci et al., 2008. Sex-specific genetic structure and social organization in
Central Asia: insights from a multi-locus study. PLoS genetics 4(9):e1000200.
Seielstad MT, Minch E, & LL. Cavalli-Sforza, 1998. Genetic evidence for a higher female migration rate in humans. Nat Genet 20(3):278-280.
Sibert A, Austerlitz F & E. Heyer, 2002. Wright-Fisher revisited: the case of fertility correlation. Theoretical Population Biology, 62, 181–197.
Verdu P, Becker NS, Froment A et al, 2013. Sociocultural behavior, sex-biased admixture, and effective population sizes in Central African Pygmies and non-Pygmies. Mol Biol Evol. 2013 Apr;30(4):918-37.
Regards connexes :
Patou-Mathis M. et C. Vercoutère, 2012. Deux regards sur l’écologie, la diversité et l’évolution des hommes du Paléolithique. Regards et débats sur la biodiversité, SFE, Regards n°39a et 39b (3 décembre 2012).
Article édité et mis en ligne par Anne Teyssèdre.
——
Bravo Evelyne pour ce regard « éclairant » (cherchez l’erreur!) sur des aspects mal connus de l’hérédité humaine, et qui met à mal bien des clichés. Et bravo pour la présentation, claire et bien illustrée…
Merci!
Merci Evelyne pour ce formidable résumé des recherches qui animent ton équipe. En quelques minutes, j’ai pu avoir un joli et clair aperçu de vos travaux marquants des 10 dernières années.
Merci!
Il est remarquable qu’Évelyne, généticienne de formation et spécialiste reconnue en la matière, articule et développe dans ses travaux l’importance de la culture dans le processus complexe d’évolution génétique de l’homme. A cet égard, je soulignerai également que, dans l’espace culturel turco-mongol non-agricole (nomade) justement, l’exogamie en tant que règle matrimoniale de la société, est présentée, et par conséquent largement enseignée, par des récits épiques dont certains comptent jusqu’à plusieurs milliers de vers. Le sujet structural de ces œuvres de la « littérature orale » est toujours la quête et la conquête par le héros principal de sa promise qui se trouve obligatoirement très-très loin de sa famille (lignée, clan, tribut) : « au-delà du troisième ciel », « sur la neuvième couche du ciel », « cachée dans le royaume souterrain » etc. On peut conclure que ces textes parlent des normes non seulement d’une exogamie clanique mais aussi d’une exogamie territoriale. Laquelle, il convient de le remarquer, a eu comme effet secondaire de promouvoir le bilinguisme.
Boris Chichlo
Merci de ces compléments.. Nous avons dans une autre étude mesurée cette exogamie géographique qui colle bien avec ces récits..
Bine cordialement
Bonjour,
et merci pour ce regard. Quand vous écrivez: « Autrement dit, on préfère se marier avec quelqu’un proche linguistiquement, même s’il est éloigné géographiquement. », est-ce bien le cas dans les populations qu’elle a étudiées, ou bien ne serait-ce pas plutôt la contraposée: « On préfère ne pas se marier avec quelqu’un qui ne parle pas la même langue, même s’il est proche géographiquement » ?
En gros, dans le premier cas, les prétendants au mariage seraient prêts à faire des centaines de kilomètres pour aller chercher un conjoint partageant leur langue, alors que dans l’autre ils choisissent juste ce qui leur convient linguistiquement dans leur environnement proche. Même si les conséquences génétiques sont à peu près les mêmes, ce n’est pas tout à fait équivalent en terme de comportement matrimonial et ça serait intéressant de savoir…
Ensuite, la question de la dichotomie entre évolution culturelle et biologique telle que présentée ici dans le cadre de la transmission du succès reproductif est un sujet bien intéressant qui mériterait bien des développements ( y aurait-il ou non des traits biologiques, tels que la force, l’agressivité ou bien encore l’intelligence des situations sociales qui favoriseraient l’accession de certains individus aux castes supérieures, et donc ipso facto à une meilleure reproduction, à tel point que beaucoup de traits dans notre biologie serait le reflet de cette sélection ?). Vaste sujet…
Finalement, je me demande s’il n’existe pas un certain nombre de contrexemples qui montrent que l’évolution génétique est souvent découplée de l’évolution linguistique, car de nombreux peuples ont changé de langue au cours de leur histoire et les déterminants linguistiques, très marqués à court terme comme le montre vos études (E. Heyer), ne sont-ils pas finalement assez labiles à l’échelle de temps plus longs ?
Bonsoir,
Merci de votre remarque sur les mariages. On ne peut pas distinguer avec nos données si on évite les mariages avec des personnes qui parlent une langue différente ou si l’on choisit préférentiellement quelqu’un qui parle une langue proche. On sait par ailleurs avec nos enquêtes ethnologiques que les gens n’hésitent pas à choisir un conjoint assez loin mais qui parle la même langue..
A l’échelle de la planète on trouve globalement une assez belle corrélation entre langue et génétique (cf. les travaux de CL Sforza), qui laisse penser que les remplacements linguistiques sont plutôt rares, bien que certains soient bien connus.
Bonjour,
Pour moi qui ne différencie pas le corps de l’esprit humain ou la transmission culturelle de la transmission biologique (ceci n’est qu’une seule et même chose, de la culture), je trouve que ce regard ne va pas encore assez loin.
Bonjour,
Evelyne H. étant partie début novembre pour plusieurs semaines en mission, elle n’a pas encore pu répondre aux commentaires et questions postées sur ce forum depuis un mois et demi. Mais elle est de retour et ne saurait tarder!
Bien cordialement à tous,
Anne