La Société Française d’Ecologie (SFE) vous propose ce regard de Barbara Demeneix, Professeur au Muséum National d’Histoire Naturelle, sur les perturbateurs endocriniens.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
———-
Perturbation endocrinienne et biodiversité :
quelques exemples
par Barbara Demeneix,
Professeur au Muséum National d’Histoire Naturelle,
Laboratoire « Evolution des Régulations Endocriniennes », UMR 7221 CNRS/MNHN,
Sorbonne Universités, Paris France.
Article édité par Anne Teyssèdre
——-
Mots clés : Perturbateurs endocriniens, hormones, pesticides, polluants industriels, physiologie animale, métamorphose, thyroïde, ecdysone, dynamique des populations, érosion de la biodiversité.
——–
- Résumé
- Quelques définitions et concepts
- Des leçons à tirer des dernières décennies
- Les organostanniques et le sexe des mollusques
- Chirurgiens bagnards, pesticides et changement climatique
- Les oiseaux migrateurs, cibles de multiples perturbateurs endocriniens
- Les chimpanzés et les pesticides
- La métamorphose des insectes et des amphibiens : une vulnérabilité particulière à la perturbation endocrinienne ?
- Le cas du glyphosate
- Conclusion et recherches futures
- Glossaire
- Bibliographie
- Forum de discussion sur ce regard
——-
Résumé
Actuellement, la prise de conscience et le débat sur les effets dits de ‘perturbation endocrinienne’ sont principalement focalisés sur la santé humaine avec l’augmentation massive de maladies non-infectieuses, telles les infertilités, les maladies métaboliques, les cancers et les maladies neuro-développementales. Cependant, les effets sur la faune sont multiples. Non seulement les démonstrations de perturbation endocrinienne chez les animaux sont très variées, mais elles ont aussi précédé les études traitant des effets néfastes sur la santé humaine, servant ainsi de système d’alerte pour l’homme.
Aujourd’hui, il faut replacer ces concepts dans une perspective écologique et planétaire, au même niveau de préoccupation que le changement climatique ou la perte de la biodiversité, sachant que la perturbation endocrinienne peut aussi être un facteur important menant à la perte de la biodiversité. Le sujet est vaste, tellement vaste qu’il faudrait plusieurs tomes pour traiter la question. Cette revue succincte propose un survol de la question avec quelques exemples choisis pour la position emblématique de l’espèce considérée ou du phénomène biologique impliqué.
Quelques définitions et concepts
La définition d’un perturbateur endocrinien la plus communément acceptée est celle d’une substance ou d’un mélange de substances, généralement mais pas exclusivement produit par synthèse industrielle, qui modifie le fonctionnement du système endocrinien et, par conséquent, induit un effet néfaste sur des individus, populations ou sous-populations.
La plupart des perturbateurs endocriniens identifiés aujourd’hui agissent en interférant avec les voies de signalisation intracellulaire contrôlées par les récepteurs nucléaires* (Casals-Casa et Desvergne, 2011 ; Préau et al., 2015). Ces derniers représentent une classe de facteurs de transcription* présente dans l’ensemble des eucaryotes* (organismes dont les cellules sont notamment dotées d’un compartiment recélant l’ADN, dit « noyau »), des levures aux mammifères. Schématiquement, ces récepteurs nucléaires (RN) sont des protéines qui se lient dans le cytoplasme* ou le noyau à certaines molécules spécifiques (dits « ligands »), le plus souvent des hormones, pour former un complexe (RN-ligand) qui se fixe sur une séquence cible de l’ADN (dite « élément de réponse »), déclenchant ou modulant la transcription de cette séquence en ARN, suivie éventuellement de sa traduction en protéine (cf. Figure 1 ci-dessous).
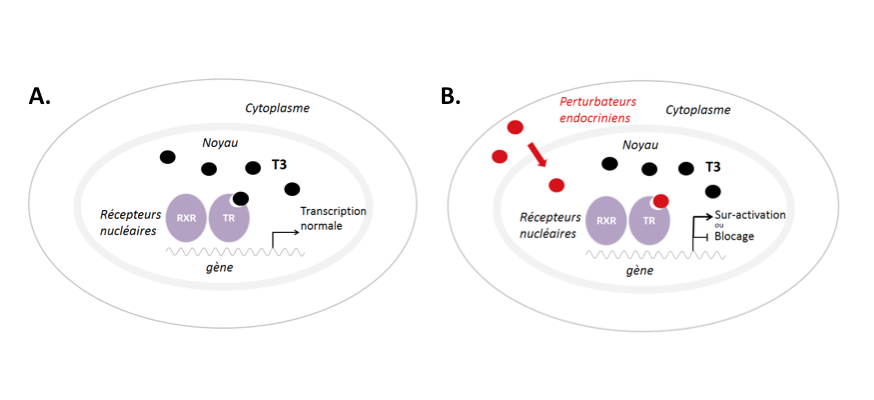
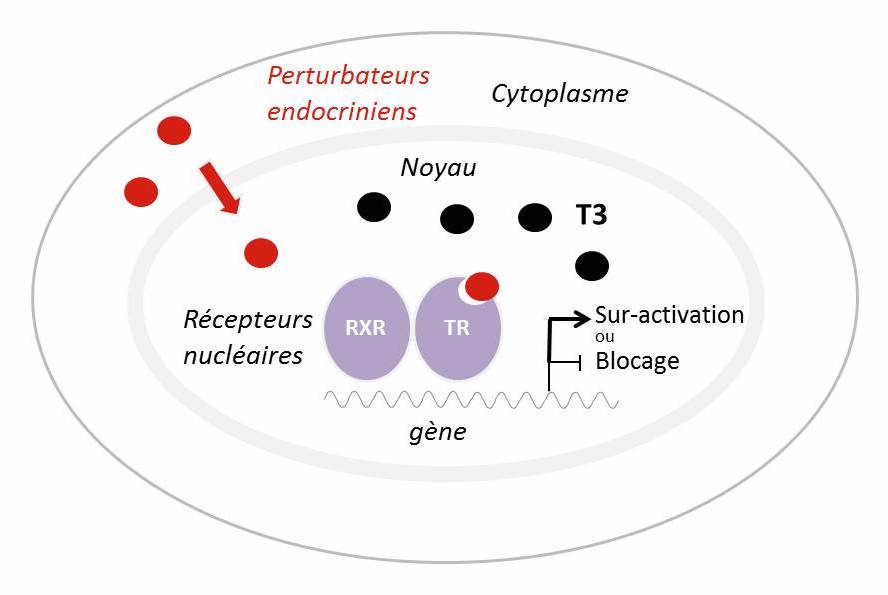
Figure 1 : Les récepteurs nucléaires comme ‘autoroutes’ entre l’environnement et la transcription des gènes
Les récepteurs nucléaires en tant que facteurs de transcription modulables par la disponibilité en hormones représentent une autoroute qui peut être empruntée par des perturbateurs endocriniens pour moduler la transcription des gènes. A : L’exemple donné représente une cellule qui répond aux hormones thyroïdiennes (T3, cercles noirs) chez un vertébré. Mais la situation peut se transposer à une cellule qui exprime des récepteurs des oestrogènes ou androgènes, voir même une cellule d’insecte qui exprime des récepteurs de l’ecdysone* avec leur parténaire hétérodimèrique l’USP (voir texte pour plus de détails). B : La transcription dans la même cellule cible peut être modulée par la présence de perturbateurs endocriniens (cercles rouges) qui brouillent les informations transcriptionelles transmises par les récepteurs nucléaires.
Une théorie propose que les récepteurs nucléaires ont évolué à partir de récepteurs de molécules xénobiotiques*, provenant du milieu extérieur (Markov et Laudet, 2011) et jouaient initialement un rôle central dans la détoxification des molécules pénétrant les cellules. L’ironie aujourd’hui est que ces mêmes récepteurs sont devenus les cibles des xénobiotiques, et surtout des perturbateurs endocriniens. En effet, leur caractéristique est d’être modulés par la disponibilité de molécules (ligands) spécifiques mais très variées – telles les rétinoïdes, les hormones sexuelles, les glucocorticoïdes, les hormones thyroïdiennes, voire même les acides gras et les sels biliaires. Activés par une grande variété de ligands, ces récepteurs sont des véhicules intracellulaires par lesquels les perturbateurs endocriniens peuvent interférer ou brouiller les informations hormonales entre les cellules.
Comme les récepteurs nucléaires sont présents dans tout le règne animal, la perturbation endocrinienne n’est pas seulement un problème de santé humaine, mais un problème global qui peut affecter des écosystèmes entiers (Carson, 1962). Pourtant, la question de la perturbation endocrinienne, au sens large, n’est pas encore suffisamment prise en compte dans les décisions réglementaires et politiques. Il est probable que la perturbation endocrinienne contribue autant que le changement climatique et la déforestation à la perte de la biodiversité massive que l’on constate aujourd’hui et représente par conséquent une bombe à retardement pour la santé des écosystèmes.
Figure 2 : Les sources de perturbateurs endocriniens. Les perturbateurs endocriniens peuvent provenir de plusieurs produits
et processus chimiques. Leur diversité explique en grande partie leur présence ubiquitaire dans l’environnement,
l’atmosphère, les milieux aquatiques et les sols.
Des leçons à tirer des dernières décennies
La première alerte pointant du doigt les effets de la perturbation endocrinienne sur la biodiversité remonte sans doute à 1962 avec la publication de Printemps Silencieux de Rachel Carson. Bien que le terme ‘perturbateur endocrinien’ n’ait pas encore été défini à l’époque, les effets néfastes sur la capacité reproductrice des populations de poissons et d’oiseaux décrits par la biologiste avaient été attribués avec une forte probabilité à l’exposition de ces populations au pesticide dichloro-diphenyl-trichloroethane (DDT).
Outre les effets sur la réduction des populations d’insectes dont certains se nourrissaient, on sait aujourd’hui que les mécanismes physiologiques mis en jeu chez les vertébrés affectés étaient bel et bien dus à la perturbation endocrinienne. En effet, il a été montré par la suite que le DDT pouvait affecter plusieurs systèmes endocriniens impliqués dans la reproduction. Une des espèces d’oiseaux les plus touchées fut le Pygargue à tête blanche (Bald eagle), l’oiseau symbole des Etats-Unis. Par la suite, on a pu démontrer qu’une des principales causes de la chute significative de la population de pygargues était la fragilisation des coquilles d’œufs et leur écrasement sous le poids des parents lors de la couvaison (Holm et al., 2006 ; Lundholm, 1997).
Or la minéralisation des œufs dépend de plusieurs systèmes endocriniens (De Matos, 2008) et peut être affectée non seulement par les pesticides en général (Hamlin et Guillette, 2010), mais aussi par de nombreux dérivés du pétrole (Rattner et al., 1984). L’attention médiatique et la prise de conscience provoquées par ce livre ont été suivies, une dizaine d’années plus tard, par la création de l’Agence pour la Protection de l’Environnement (EPA) aux Etats-Unis et une interdiction d’utiliser du DTT aux Etats-Unis en 1972. Cependant, la Convention de Stockholm recommande encore son usage dans certains cas, notamment dans le cadre de la lutte contre le paludisme en Afrique du Sud, et ceci en dépit des effets néfastes sur la santé humaine (Rogan et Chen, 2005) et la biodiversité.
Déjà en 1962, de manière presciente, Rachel Carson avait prédit que les effets sur la biodiversité allaient tôt ou tard se répercuter sur les populations humaines. L’ironie veut qu’il ait fallu près de cinquante ans pour être en mesure de bien le démontrer. Effectivement, ce n’est qu’en 2015 que, grâce à l’analyse de centaines d’échantillons de sang provenant du cordon ombilical collecté dans les années 1950, Barbara Cohn et son équipe ont pu montrer un risque accru du cancer du sein chez les filles exposées lors de la période prénatale à un taux élevé de DDT (Cohn et al., 2015). L’ironie réside également dans le fait qu’aujourd’hui, la situation s’est quelque peu inversée, par rapport aux prédictions de Rachel Carson, dans le sens où l’on s’inquiète davantage des effets de la perturbation endocrinienne sur la santé des populations humaines, alors que les effets sur la biodiversité sont tout aussi importants, voire encore plus dévastateurs.
En outre, bien que les Etats-Unis aient été parmi les premières nations à introduire des législations et des suivis détaillés relatifs à l’utilisation des pesticides, on assiste aujourd’hui à une dérégulation massive outre-Atlantique. Un exemple est la ré-autorisation aux Etats-Unis et ailleurs du chlorpyrifos, pesticide organo-phosphate dont on a pourtant démontré qu’il augmentait le risque de maladies neuro-développementales telles les maladies du spectre autistique (Shelton et al., 2014).
Paradoxalement, en dépit du fait que le DDT ait été interdit aux Etats-Unis dans les années 1970, les données du Programme des Nations Unis pour l’Environnement montrent que la production chimique mondiale (chiffrée en termes de valeur économique) a été multipliée par 300 depuis cette date, et que la plupart de ces produits chimiques n’ont pas été testés pour leurs effets toxiques ni pour leurs effets sur la fonction endocrinienne.
On associe rarement la perte de biodiversité à la perturbation endocrinienne, la liant généralement davantage à la pollution chimique et au changement climatique. Ceci est bien démontré dans le contexte de ‘One Health’, expression pour laquelle même l’Institut National de la Recherche Agronomique (INRA) n’a pas encore trouvé de traduction satisfaisante. En effet, sous ce chapeau « d’une seule santé », initié par le Wildlife Conservation Society, on trouve regroupées plusieurs organisations et plusieurs idées-principes dont la santé animale, la santé humaine et le respect des écosystèmes avec un leitmotiv qui vise à affirmer « le partage des responsabilités et la coordination des actions globales pour gérer les risques sanitaires aux interfaces animal-homme-écosystèmes ».
L’accent est donc mis sur les risques sanitaires pour l’homme, notamment les risques d’épidémies (par exemple de grippe aviaire). De plus, quand on se penche sur les mots-clefs associés, on trouve bien le changement climatique, la résistance aux antibiotiques et l’intensification de l’agriculture, mais jamais la perturbation endocrinienne.
Pourtant, plusieurs exemples bien documentés montrent les effets de perturbation endocrinienne affectant chaque groupe d’animaux et chaque système endocrinien, le tout étant intrinsèquement lié à l’évolution des régulations endocriniennes [(Markov et al., 2009). Dans les exemples qui ont été sélectionné ci-dessous, l’objectif a été de souligner les mécanismes communs et surtout l’implication des récepteurs nucléaires comme cibles des perturbateurs endocriniens.
Les organostanniques et le sexe des mollusques
Outre le DDT, plusieurs produits chimiques ont été rapidement identifiés comme perturbateurs endocriniens agissant sur les capacités de reproduction ou d’adaptation de plusieurs groupes d’animaux. Parmi les multiples exemples, on peut citer le cas d’un organostannique*, le tributylétain (TBT), un composant des peintures anti-salissures appliquées sur la partie inférieure des bateaux pour empêcher le développement des organismes qui s’y attachent. Son utilisation a été associée à la chute de la production aquacole de plusieurs espèces de mollusques, dont les coquilles Saint-Jacques, les moules et surtout les huitres. On a pu mettre en évidence plusieurs cas de masculinisation de femelles mollusques, dont des gastéropodes (le Bulot en particulier), qui montraient une sensibilité extrême à cette molécule (TBT) avec une masculinisation (« imposex ») et infertilité des femelles à partir de très faibles concentrations, de quelques nanogrammes par litre.
La réduction massive de production des huitres dans la baie d’Arcachon a poussé la France à être le premier pays à interdire ce composé en 1982, suivi par d’autres pays Européens et par une interdiction internationale en 1999. Aujourd’hui, on sait que le TBT agit, comme beaucoup d’autres perturbateurs endocriniens, en interférant avec les récepteurs nucléaires (RN) de la classe des récepteurs aux rétinoïdes (RXR) et les récepteurs activés par les proliférateurs des peroxysomes* (PPAR) (Nakanishi, 2008).
Souvent les récepteurs nucléaires agissent en couple. Par exemple, les RXR cités ci-dessus sont des partenaires des récepteurs des PPAR, mais aussi des récepteurs des hormones thyroïdiennes (TR), de l’acide rétinoïque (RAR) et de bien d’autres molécules. En tant que partenaire des TR, le TBT a été montré comme perturbateur endocrinien de plusieurs mécanismes et régulations physiologiques TR/RXR dépendantes, allant des régulations métaboliques centrales, au niveau hypothalamique (Santos-Silva et al., 2017) ou au niveau des cellules musculaires, comme démontré récemment chez la carpe (Li et al., 2016).
Chirurgiens bagnards, pesticides et changement climatique
Un autre processus dépendant des hormones thyroïdiennes et cible des perturbateurs endocriniens est le recrutement, au niveau des récifs coralliens, des larves pélagiques de poissons dont le stade adulte est récifal, tel le Chirurgien bagnard Acanthurus triostegus.
Comme pour la métamorphose des amphibiens (Tata, 1998) et celle des poissons plats (Shreiber, 2013) tels le Turbot et la Sole, le développement des « chirurgiens bagnards » est dépendant d’un pic d’hormones thyroïdiennes. Le changement climatique, l’acidification des océans, ainsi que la surexploitation des ressources, présentent les menaces principales des récifs coralliens. A ceci s’ajoute aussi la pollution chimique, notamment les pesticides dont plusieurs sont des perturbateurs endocriniens avérés.
Très récemment, on a démontré les effets néfastes d’un pesticide organo-phosphate, le chlorpyrifos, chez le Chirurgien bagnard (Holzer et al., 2017). La publication montre non seulement que les hormones thyroïdiennes sont nécessaires pour le recrutement de plusieurs espèces de larves pélagiques de poissons associées aux bancs de coraux, mais aussi que le chlorpyrifos interfère avec le développement de l’intestin adulte et la capacité du poisson de se nourrir d’algues sur le corail. Tous ces changements physiologiques et comportementaux dépendent des hormones thyroïdiennes. Ce pesticide est déjà connu pour ses effets de perturbation thyroïdienne chez des rongeurs. Les hormones thyroïdiennes étant nécessaires pour le développement du cerveau chez les mammifères, dont l’homme, il n’est pas étonnant de voir que des études épidémiologiques ont associé ce pesticide à une baisse de QI et une augmentation du risque de troubles du spectre autiste chez les enfants de femmes fortement exposées pendant leur grossesse.
Les oiseaux migrateurs, cibles de multiples perturbateurs endocriniens
Un des exemples les plus frappants illustrant les effets des perturbateurs endocriniens (PE) concerne les oiseaux migrateurs. Plusieurs régulations physiologiques des oiseaux migrateurs sont sous le contrôle du système endocrinien, et particulièrement des hormones thyroïdiennes. Les adaptations saisonnières régulent évidement la reproduction, mais aussi le métabolisme, les modifications de structure musculaire, la prise de poids en préparation de la migration, ainsi que le contrôle cérébral des mécanismes de navigation mis en jeu. Dans tous ces changements physiologiques subtils nécessaires à la migration, l’hormone thyroïdienne est essentielle. Un grand champion de l’endurance est la Barge rousse, un échassier grand migrateur qui vole d’une traite de l’Arctique jusqu’en Tasmanie ou en Nouvelle-Zélande à la fin de l’été de l’hémisphère Nord, puis revient au début du printemps (lorsque les saisons s’inversent).
Christy Morrissey, de l’Université de la Saskatchewan, étudie quant à elle un oiseau migrateur bien plus connu, l’Etourneau sansonnet (Sturnus vulgaris). Elle a montré qu’une catégorie de PE affectant les hormones thyroïdiennes, les polychlorés bisphénols (PCB), interfère avec les adaptations métaboliques nécessaires pour la migration. Comme le DDT, les PCB ont été bannis dans les années 1970, mais comme ils sont très persistants, les êtres humains, la faune et la flore y sont toujours exposés à des quantités non-négligeables. Malheureusement, les mêmes perturbateurs thyroïdiens endommagent ces mécanismes neuronaux complexes nécessaires à l’orientation dès le début de la migration. Les oiseaux sont donc confrontés à la perturbation, non seulement de leur système de navigation, mais ils subissent aussi une altération de leur capacité métabolique (Zahara et al., 2015 ; Flahr et al., 2015).
Les chimpanzés et les pesticides
Un autre exemple, relatif aux primates, concerne un projet que notre équipe conduit en collaboration avec Sabrina Krief, primatologue au Muséum National d’Histoire Naturelle à Paris. Avec ses collègues de l’Université Makerere, en Ouganda, Sabrina Krief étudie les effets d’une surutilisation des pesticides sur les populations de chimpanzés et de babouins. Compte tenu de la sensibilité de ces groupes de chimpanzés à l’intervention humaine, Sabrina et ses collègues doivent recourir à des méthodes strictement non-interventionnistes pour obtenir des échantillons d’excréments et d’urine. Elle a déjà observé des malformations faciales prononcées chez les chimpanzés qui se nourrissent de maïs cultivé.
Notre hypothèse de départ rassemble les deux idées selon lesquelles une grave carence en hormones thyroïdiennes, durant le développement du fœtus, affecte la forme de la face (comme chez les crétins) et que beaucoup des pesticides utilisés sont des perturbateurs thyroïdiens reconnus. Le rôle de l’équipe des endocrinologues consiste à analyser les niveaux d’hormones thyroïdiennes dans l’urine des chimpanzés et à établir les associations potentielles entre l’exposition aux pesticides et les malformations cranio-faciales. Nous avons pu montrer que des taux élevés de plusieurs pesticides sont présents dans le maïs cultivé à proximité du parc (Krief et al., 2017), sachant que plusieurs de ces pesticides affectent la signalisation par les hormones thyroïdiennes.
Les effets des pesticides sur la structure faciale des chimpanzés peuvent peut-être paraître moins importants que d’autres répercussions sur la biodiversité. Cependant, comme cela a été démontré avec la thalidomide, il est plus facile de prendre conscience des malformations des membres et des faces que d’autres formes d’affections moins visibles, touchant autant ou davantage des capacités adaptives. En outre, dans le cas des malformations des chimpanzés, il s’agit d’un des rares effets que nous puissions mesurer de façon éthique chez ces espèces sentinelles et emblématiques, sans les déranger.
La métamorphose des insectes et des amphibiens : une vulnérabilité particulière à la perturbation endocrinienne ?
On constate actuellement une perte massive de la biodiversité, certaines espèces étant plus affectées que d’autres. Différentes études montrent notamment que les populations d’insectes (Hallmann et al., 2017) et d’amphibiens (Collins, 2010) sont particulièrement touchées. Or, bien que les mécanismes endocriniens qui régulent la métamorphose des amphibiens (Fig. 3A) et celle des autres vertébrés et des insectes (Fig. 3B) diffèrent dans les détails, ils partagent tout de même plusieurs aspects. Le premier est que la régulation endocrinienne implique dans les deux cas des récepteurs nucléaires. Pour les insectes, il s’agit d’un hétéro-dimère entre le récepteur de l’ecdysone et l’ultraspiracle (USP), l’homologue des RXR chez les mollusques et les vertébrés (Markov et Laudet, 2011).
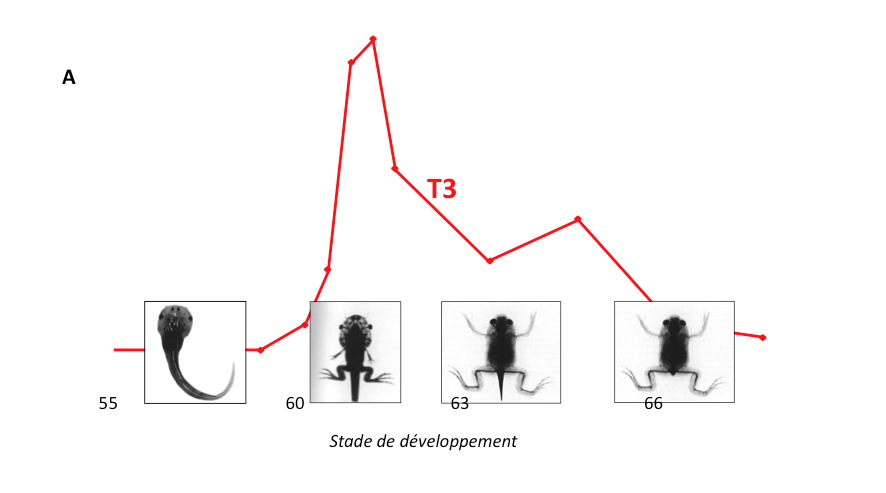
Figure 3 : Les transitions développementales régulées par les hormones thyroïdiennes chez des vertébrés, ou par l’écdysone (hormone déclenchant la métamorphose) chez les insectes, peuvent être la cible de perturbateurs endocriniens.
A: La métamorphose des amphibiens (tout comme celle des poissons plats) est régulée par les hormones thyröidiennes. La métamorphose est associée à un remaniement épigénétique*. Comme les hormones thyröidiennes sont des cibles très vulnérables à la perturbation endocrinienne, les amphibiens sont aussi très sensibles aux perturbateurs endocriniens
présents dans l’eau,l’atmosphère ou le sol. Stade de développement: nombre de jours.
T3: Concentration sanguine d’hormone thyroïde T3.
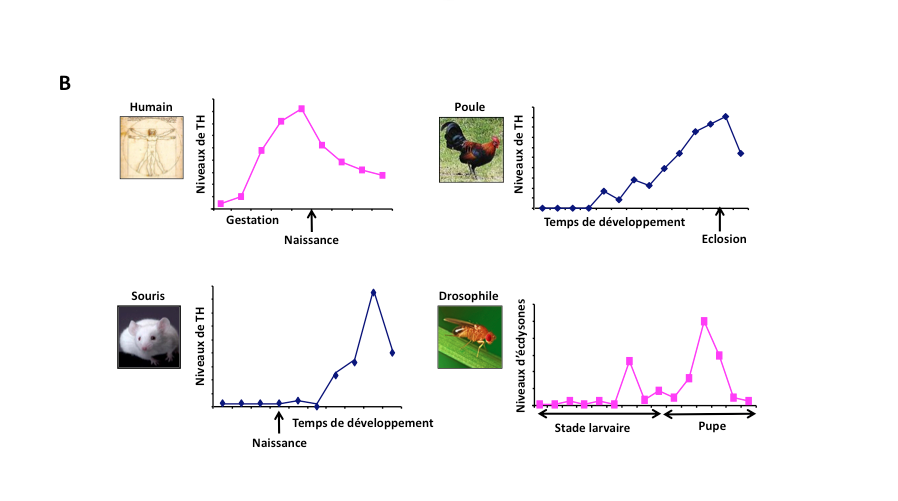
B: Les hormones thyroïdiennes orchestrent les transitions développementales critiques chez tous les vertébrés.
De plus, leurs récepteurs sont homologues des récepteurs de l’écdysone qui régulent la métamorphose des insectes,
cibles aussi des perturbateurs endocriniens.
Un autre facteur pouvant expliquer la sensibilité particulière des animaux qui subissent des métamorphoses est le fait que, chez de nombreuses espèces, les individus sont exposés à une certaine catégorie de polluants (par exemple, dans l’eau) au stade larvaire, puis à d’autres polluants (par exemple, dans l’air) au stade adulte.
Un autre aspect, non pris en compte encore dans la législation, est celui des modifications épigénétiques* qui peuvent être conservées par la suite. Un exemple : la métamorphose qui dépend des hormones thyroïdiennes chez les poissons téléostéens et les amphibiens. Ce même pic d’hormones thyroïdiennes est caractéristique des transitions développementales chez tous les vertébrés (Laudet, 2011) et représente une suite de modifications épigénétiques* dans chaque espèce. Les hormones thyroïdiennes induisent des enzymes modulatrices de l’état de méthylation de l’ADN, un exemple de régulation épigénétique. Comment les polluants et les perturbateurs endocriniens affectent ces régulations reste une question encore peu étudiée.
Le cas du glyphosate
Actuellement, en France et au niveau européen, le cas du glyphosate est beaucoup discuté. On entend souvent dire que cet herbicide ne peut pas être un perturbateur endocrinien parce que l’enzyme ciblée n’est pas exprimée par les cellules animales – n’étant présente que chez certaines plantes et micro-organismes. Effectivement, l’enzyme ciblée par le glyphosate est le 5-enolpyruvylshikimate-3-phosphate synthétase, nécessaire à la production des acides aminés et des produits secondaires chez plusieurs plantes et microorganismes. Cependant, ce serait oublier a minima trois facteurs importants.
Premièrement, les enzymes de la flore intestinale des vertébrés et celle des insectes expriment cette enzyme et peuvent être affectées négativement pas l’herbicide (Vandenberg et al., 2017 ; Shebata et al., 2013, Ackermann et al., 2015 ; Myers et al., 2016). Deuxièmement, le glyphosate n’est jamais appliqué seul, mais avec des adjuvants, le plus souvent sous forme de Roundup ®, produit qui a souvent été démontré comme perturbateur endocrinien (pour revue, voir Vandenberg et al., 2017).
Le troisième facteur à prendre en compte, bien que non directement relié à des effets de perturbation endocrinienne, est évidemment les effets du glyphosate sur la fertilité des sols, soit en formant des complexes avec des minéraux (i.e., par action de chélation*), soit par action sur les bactéries bénéfiques, y compris celles qui fixent l’azote (N) (Vandenberg et al., 2017 ; Myers et al., 2016 ; Van Bruggen et al., 2017).
Conclusion et futures recherches
Cette revue présente quelques exemples d’actions de perturbateurs endocriniens illustrant la diversité des organismes affectés, dans l’ensemble du règne animal et au-delà. Les exemples cités sont pour la plupart des effets de molécules individuelles, mais on sait aujourd’hui que tout être vivant est exposé à des mélanges complexes, composés de plusieurs dizaines voire centaines de molécules aux effets synergiques potentiellement démultipliés.
La recherche future doit prendre en compte cette complexité moléculaire, tout en embrassant les concepts d’interactions entre espèces, la possibilité de régulations épigénétiques* et éventuellement transgénérationnelles.
Glossaire
- Chélation : Formation physico-chimique d’un complexe impliquant des liaisons chimiques entre un élément métallique et une autre molécule. La chélation limite la disponibilité du métal pour entrer, par exemple, dans une réaction biologique.
- Epigénétiques (modifications, régulations) : Modifications de réponses génétiques résultant de modifications de la transcription de séquences d’ADN, transmises à la descendance sur une ou plusieurs générations, sans mutation de ces séquences.
- Facteurs de transcription : Protéines qui se fixent à l’ADN et modifient la transcription de gènes cibles.
- Organostannique : Molécule organique contenant du carbone et de l’étain.
- Peroxysome : organite intracellulaire entouré par une membrane simple et ne contenant pas de matériel génétique. Les récepteurs nucléaires ‘PPAR’ ont été ainsi appelés car leur activation provoque la prolifération des peroxysomes.
- Récepteurs nucléaires : Grande classe de facteurs de transcription dont l’activité est le plus souvent modulée par des hormones tels les stéroïdes, les hormones thyroïdiennes, les rétinoïdes et les glucocorticoïdes (hormones de stress).
- Xénobiotique : Molécule ou composé chimique d’origine extérieure, non produit par l’organisme concerné.
Bibliographie
Ackermann W. et al., 2015. The influence of glyphosate on the microbiota and production of botulinum neurotoxin during ruminal fermentation. Curr Microbiol, 70(3): p. 374-82.
Carson, R., L. Darling, and L. Darling, 1962. Silent spring. Boston Cambridge, Mass.: Houghton Mifflin ; Riverside Press. x, 368 p.
Casals-Casas C. et Desvergne B., 2011. Endocrine disruptors: from endocrine to metabolic disruption. Annu Rev Physiol, 73: p. 135-62.
Cohn B.A. et al., 2015. DDT Exposure in Utero and Breast Cancer. J Clin Endocrinol Metab, 100(8): p. 2865-72.
Collins J.P., 2010. Amphibian decline and extinction: what we know and what we need to learn. Dis Aquat Organ, 92(2-3): p. 93-9.
De Matos R., 2008. Calcium metabolism in birds. Vet Clin North Am Exot Anim Pract, 11(1): p. 59-82, vi.
Flahr L.M. et al., 2015. Developmental Exposure to Aroclor 1254 Alters Migratory Behavior in Juvenile European Starlings (Sturnus vulgaris). Environ Sci Technol, 49(10): p. 6274-83.
Hallmann C.A. et al., More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One, 12(10): p. e0185809.
Hamlin H.J. et Guillette L.J, Jr., 2010. Birth defects in wildlife: the role of environmental contaminants as inducers of reproductive and developmental dysfunction. Syst Biol Reprod Med, 56(2): p. 113-21.
Holm L. et al., 2006. Embryonic exposure to o,p’-DDT causes eggshell thinning and altered shell gland carbonic anhydrase expression in the domestic hen. Environ Toxicol Chem, 25(10): p. 2787-93.
Holzer G. et al., 2017. Fish larval recruitment to reefs is a thyroid hormone-mediated metamorphosis sensitive to the pesticide chlorpyrifos. Elife, 6.
Krief S. et al., 2017. Agricultural expansion as risk to endangered wildlife: Pesticide exposure in wild chimpanzees and baboons displaying facial dysplasia. Sci Total Environ, 598: p. 647-656.
Laudet V., 2011. The origins and evolution of vertebrate metamorphosis. Curr Biol, 21(18): p. R726-37.
Li Z.H., Li P., et Shi Z.C, 2016. Chronic effects of tributyltin on multiple biomarkers responses in juvenile common carp, Cyprinus carpio. Environ Toxicol, 31(8): p. 937-44.
Lundholm C.D., 1997. DDE-induced eggshell thinning in birds: effects of p,p’-DDE on the calcium and prostaglandin metabolism of the eggshell gland. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol, 118(2): p. 113-28.
Markov G.V. et al., 2009. Independent elaboration of steroid hormone signaling pathways in metazoans. Proc Natl Acad Sci U S A, 106(29): p. 11913-8.
Markov G.V. et Laudet V., 2011. Origin and evolution of the ligand-binding ability of nuclear receptors. Mol Cell Endocrinol, 334(1-2): p. 21-30.
Myers J.P. et al., 2016. Concerns over use of glyphosate-based herbicides and risks associated with exposures: a consensus statement. Environ Health, 15: p. 19.
Nakanishi T., 2008. Endocrine disruption induced by organotin compounds; organotins function as a powerful agonist for nuclear receptors rather than an aromatase inhibitor. J Toxicol Sci, 33(3): p. 269-76.
Préau L. et al., 2015. Thyroid hormone signaling during early neurogenesis and its significance as a vulnerable window for endocrine disruption. Biochim Biophys Acta, 1849(2): 112-21.
Rattner B.A. et al., 1984. Avian endocrine responses to environmental pollutants. J Exp Zool, 232(3): p. 683-9.
Rogan W.J. et Chen A., 2005. Health risks and benefits of bis(4-chlorophenyl)-1,1,1-trichloroethane (DDT). Lancet, 366(9487): p. 763-73.
Santos-Silva A.P. et al., 2017. Frontiers in endocrine disruption: Impacts of organotin on the hypothalamus-pituitary-thyroid axis. Mol Cell Endocrinol.
Schreiber A.M., 2013. Flatfish: an asymmetric perspective on metamorphosis. Curr Top Dev Biol, 103: p. 167-94.
Shehata A.A. et al., 2013. The effect of glyphosate on potential pathogens and beneficial members of poultry microbiota in vitro. Curr Microbiol, 66(4): p. 350-8.
Shelton J.F. et al., 2014. Neurodevelopmental disorders and prenatal residential proximity to agricultural pesticides: the CHARGE study. Environ Health Perspect, 122(10): p. 1103-9.
Tata J.R., 1998. Amphibian metamorphosis as a model for studying the developmental actions of thyroid hormone. Cell Res, 8(4): p. 259-72.
Van Bruggen A.H.C. et al., 2017. Environmental and health effects of the herbicide glyphosate. Sci Total Environ, 616-617: p. 255-268.
Vandenberg L.N. et al., 2017. Is it time to reassess current safety standards for glyphosate-based herbicides? J Epidemiol Community Health, 71(6): p. 613-618.
Zahara A.R. et al., 2015. Latent cognitive effects from low-level polychlorinated biphenyl exposure in juvenile European starlings (Sturnus vulgaris). Environ Toxicol Chem, 34(11): p. 2513-22.
.
Regard connexe :
Dussud C. et J-F. Ghiglione, 2014. La dégradation des plastiques en mer. Regards et débats sur la biodiversité, SFE, Regard n°63 (26 décembre 2014).
Article édité et mis en ligne par Anne Teyssèdre.
——
Très bien écrit, merci Mme Domeneix. Une petite remarque et une petite question. La remarque concerne Acanthurus triostegus qui est le chirurgien bagnard et non le poisson clown, mais ça ne change rien au schmilblick. Et la question concerne le glyphosate: quels sont ces perturbateurs contenus dans les adjuvants du Round-up ? On en parle souvent mais apparemment leur nature est peu connue, est-ce tabou ?
Merci beaucoup !
Bonjour,
il me semble que le formaldéhyde est un des adjuvants les plus toxiques, dans sa forme originale ou de décomposition… Mais il en existe beaucoup d’autres, cf les travaux du CRIIGEN.
Merci! je vais regarder les données sur le formaldehyde.
En réponse à Amphiprion :
Vous avez tout à fait raison, pour le nom français d’A. triostegus: c’est bien le chirurgien bagnard. Désolée pour l’erreur! je vais demander de corriger le texte.
[C’est fait : erreur corrigée (le 9 février) dans le texte en ligne! Anne T. ]
Pour le round up : Plusieurs jeux de données montre les effets de perturbation endocrinienne du round up – dont les actions sur l’axe thyroidien :Toxicology. 2017 Feb 15;377:25-37. doi: 10.1016/j.tox.2016.11.005
Excellent regard, portant sur un des problèmes majeurs à l’heure actuelle. Espérons que ce cri d’alarme sera (enfin) entendu. Moi aussi, j’aimerais en savoir plus sur les adjuvants du glyphosate contenus en particulier dans le RoundUp. J’ai eu récemment à évaluer un article scientifique (provenant de l’équipe de chercheurs de Monsanto) concernant les effets du glyphosate sur différents groupes animaux du sol. J’ai émis la remarque que ce genre d’étude était parfaitement inutile car le glyphosate n’est jamais utilisé seul mais lié à des produits destinés à faciliter sa pénétration dans la cellule et éventuellement à augmenter ses effets. Bien entendu, ma remarque a été évacuée, car seule la molécule active fait l’objet d’une réglementation (comme pour les médicaments), les adjuvants relevant du secret de fabrication. L’article a été publié, malgré mes remarques négatives…
Quel dommage! Vous avez tout à fait raison, le glyphosate n’est jamais utilisé seul, plusieurs publications démontrent des effets plus néfastes en formulation.
Merci pour cet article alarmant mais intéressant, que je peux comprendre (sauf quelques lignes du début) alors que je suis pas scientifique. Je ne vois pas ce que je peux faire en tant qu’individu pour lutter contre ce fléau grandissant de l’empoisonnement de la vie terrestre, mais j’aurais un argument de plus pour contrer ceux qui osent encore utiliser du Roundup. Les Regards de la SFE devraient faire l’objet d’une publication grand public tant ils sont passionnants.
Merci pour votre avis!
Bonjour,
Oui je rejoins Marie-Noelle. J’ai quitté le milieu scientifique actif pour participer au « redressement » (sourire). J’observe la SFE depuis quelques années. Cette association est un excellent signe au sein de la communauté scientifique. Cette vitrine nous montre aussi des difficultés propre à la communauté. La prise de position sur les publications est un acte que je trouve extrêmement bénéfique. Je galère depuis toujours pour accéder à l’information (prix, réservée, etc).
En tant qu’individus nous pouvons pendre position et agir conformément à notre prise de position, même si l’on de comprends pas tout. Par exemple choisir un polluant facilement identifiable dans notre vie quotidienne et le bannir. Assurément notre entourage réagira et cela aura un effet boule de neige, voire avalanche. Je ne vous donne pas d’exemple car vous constaterez que le choix est très difficile. Cet exercice nous met en situation de pollueur consciencieux et finalement bien embêté quand il s’agit d’agir de façon radicale. Pourtant c’est faisable.
Par ailleurs, lutter contre un système de pensées est rarement le choix le plus facile. L’idée de conforter, de renforcer un système différent et plus adapté, qui trace son propre chemin, est bien plus porteuse je trouve. Regardez Slow Food, Le bio, WWF, Les vins naturels, SFE etc. Alors je vote, j’adhère et j’encourage (clap clap). Hip… Bravo à Mme Demeneix et son équipe!
Merci,
Franck
Une superbe analyse sur les perturbateurs endocriniens. N’étant pas scientifique, vos recherches me semblent sensées et très utiles.
La vérité est hélas étouffée par des intérêts économiques. L’économie mondiale basée sur la compétitivité du moins cher use encore plus le vivant. L’être humain aussi, bien entendu. Il faut médiatiser encore plus vos recherches et rester propre, chose dont je ne doute pas une seconde. Merci de nous dévoiler ces faits,et je vous souhaite une bonne continuation.