La Société Française d’Ecologie (SFE) vous propose ce regard de Sébastien Barot, Directeur de recherche à l’IRD, sur l’évolution et les conséquences de la compétition dans divers domaines.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
———-
La compétition, l’évolution,
le système de recherche et la société
par Sébastien Barot,
Directeur de recherche en écologie à l’IRD, Vice-Président de la SFE
——-
Mots clés : Compétition, écologie, évolution, recherche, société.
——-
- Importance des relations de compétition en écologie
- Compétition, évolution et optimisation
- Application au système de recherche scientifique
- Application aux sociétés humaines
- Conclusion
- Glossaire
- Bibliographie
- Forum de discussion sur ce regard
——-
Importance des relations de compétition en écologie
Les organismes sont liés par différents types d’interactions : compétition, prédation, mutualisme… La compétition est une des interactions écologiques les plus étudiées. Elle est très souvent liée à la consommation d’une ressource commune. Pour vivre, grandir, se défendre ou se reproduire, tous les organismes utilisent des ressources (de l’énergie et de la matière) qu’ils puisent dans l’environnement. Les producteurs primaires (par exemple les plantes) consomment de l’eau, du CO2, des nutriments minéraux et utilisent la lumière comme source d’énergie pour fabriquer leur matière organique. Les animaux consomment des producteurs primaires (cas des herbivores), ou d’autres animaux (cas des carnivores). Les organismes vivants sont aussi en compétition pour l’habitat ou l’espace qu’ils occupent.
Les conséquences de la compétition sont multiples et se font sentir à des échelles écologiques très diverses. Le mécanisme fondamental est qu’une ressource consommée par un individu, ou par une population* donnée, n’est plus disponible pour les autres individus ou pour les autres populations. De ce fait, toute chose égale par ailleurs, si un individu parvient à accaparer plus de ressources que ses compétiteurs, il va grandir plus vite, vivre plus longtemps et/ou avoir plus de descendants. Si une population d’une espèce arrive à augmenter la part des ressources qu’elle peut exploiter, la biomasse totale de cette espèce devrait augmenter, ainsi que le nombre de ses individus. Ce type de raisonnement a conduit au principe d’exclusion compétitive (Hardin 1960) qui suggère que quand deux espèces ont une niche écologique* très proche, et sont donc en compétition pour la même ressource limitante, l’espèce qui exploite au mieux cette ressource éliminera l’autre, au moins sur le long terme. Tilman (1982) a développé une théorie légèrement différente suggérant que l’espèce qui est capable d’abaisser le plus la disponibilité d’une ressource, et d’en tirer parti, élimine toutes ses concurrentes.
D’une manière générale, la quantité totale des ressources et la compétition pour les ressources considérées comme limitantes, régulent la démographie des espèces si bien que leurs populations ne peuvent pas croître indéfiniment. Cela constitue peut-être une des lois universelles de l’écologie (Turchin 2001). Ce type de raisonnement se rapproche aussi fortement de certaines intuitions de Malthus (1798). Utilisant des modèles mathématiques simples, ce dernier a montré dès la fin du 18e siècle qu’une croissance démographique exponentielle des populations humaines ne peut être soutenue indéfiniment, parce que les ressources utilisées par les humains sont finies ne peuvent croitre aussi vite. Cela devait alors conduire à des catastrophes démographiques.
L’impact écologique de la compétition pour les ressources se poursuit à l’échelle évolutive (au sens darwinien). La sélection naturelle* opère car les ressources sont limitantes. Ainsi chaque individu ne peut se procurer qu’une quantité finie de ressources, et transformer celle-ci en une probabilité de survie et en un nombre de descendants finis. Cela est dû à la fois à la limitation par les ressources et à la compétition, et au fait que chaque individu doit allouer la quantité de ressource finie qu’il a acquis à des fonctions très diverses (Stearns 1992): acquisition des ressources, métabolisme de base, locomotion, croissance, reproduction, protection contre différents prédateurs et parasites, plasticité permettant de répondre à la variabilité environnementale…
Ces processus conduisent au fait que les individus ayant la meilleure stratégie d’allocation des ressources ont plus de descendants et que les gènes qui déterminent cette stratégie peuvent se répandre et devenir majoritaires au sein d’une espèce. Les individus ayant une moins bonne stratégie ont une survie plus faible et moins de descendants. Leur stratégie et les gènes correspondants vont avoir tendance à disparaitre. La compétition explique aussi l’adaptation à des conditions environnementales changeantes. De nouvelles conditions modifient la stratégie d’allocation permettant d’acquérir au mieux les ressources et d’avoir le plus de descendants. Sans limitation par les ressources (ressource abiotique* pour les producteurs primaires, aliment, habitat…), chaque individu pourrait théoriquement avoir une descendance infinie et la sélection naturelle n’aurait plus de prise.
Compétition, évolution et optimisation
La situation présentée jusqu’ici correspond jusqu’à un certain point à une caricature du message darwinien : les « plus forts » acquièrent plus de ressources, leur permettant d’éliminer les « plus faibles », le tout étant le moteur de l’évolution qui conduit à l’avènement des « plus forts ». Cette vision n’est pas entièrement fausse et permet de comprendre, par exemple, comment la sélection naturelle a permis la colonisation de la plupart des milieux terrestres et aquatiques (y compris des environnements extrêmement inhospitaliers, cas des bactéries extrêmophiles vivant dans des milieux très acides ou très chauds) par les êtres vivants et la diversification de leurs niches écologiques.
On peut à juste titre s’émerveiller de l’incroyable diversité des inventions de l’évolution. Cependant, ces processus n’ont pas de raison particulière d’optimiser des propriétés collectives des populations ou des écosystèmes, comme la production primaire. Clairement, le critère impliqué par la sélection naturelle est la transmission des gènes par un individu à ses descendants. Cette transmission peut éventuellement être liée à la capacité de cet individu à produire de la biomasse mais n’a pas de raison d’être liée à la biomasse collective de l’espèce, sauf dans des cas particuliers. Cela signifie qu’au cours de l’évolution la biomasse d’une espèce peut diminuer, ce qui dans des cas extrêmes peut fragiliser une espèce jusqu’à la conduire à l’extinction. On peut alors parler de suicide évolutif (Parvinen 2005, Boudsocq et al. 2011).
Un problème de l’évolution sous l’effet de la compétition est qu’elle peut dans certains cas exacerber la compétition et déboucher sur « une tragédie des biens communs » (Hardin 1968). Quand une ressource est partagée par tous les individus d’une population ou toutes les espèces en présence, l’évolution a tendance à favoriser les individus qui consomment le plus vite cette ressource, ce qui leur permet d’empêcher les autres individus/espèces d’utiliser cette ressource. Cela peut conduire à des stratégies sub-optimales du point de vue de la biomasse totale produite par la population ou la communauté. Cela peut aussi conduire à l’épuisement de la ressource et à la disparition des espèces dépendant de cette ressource (forme de suicide évolutif) : la compétition et l’évolution peuvent compromettre la durabilité d’un système écologique.
Par exemple, il a été suggéré que les plantes peuvent investir énormément dans leur système racinaire et leur surface foliaire pour ne pas perdre la compétition pour les nutriments minéraux et la lumière (Gersani et al. 2001, Denison et al. 2010). Cela constitue une forme de « course aux armements ». Cet investissement est souvent bien trop élevé par rapport à ce qui serait strictement nécessaire, et réduit la production de biomasse (voir Figure 1). Autrement dit, si une plante augmente sa surface foliaire, elle peut produire plus de biomasse, mais si tous les individus d’une population augmentent leur surface foliaire la compétition entre individus augmente, chaque individu faisant de plus en plus d’ombre à ces voisins. La quantité de matière organique fabriquée par cm2 de surface foliaire diminue parce que la quantité de lumière incidente n’augmente pas.
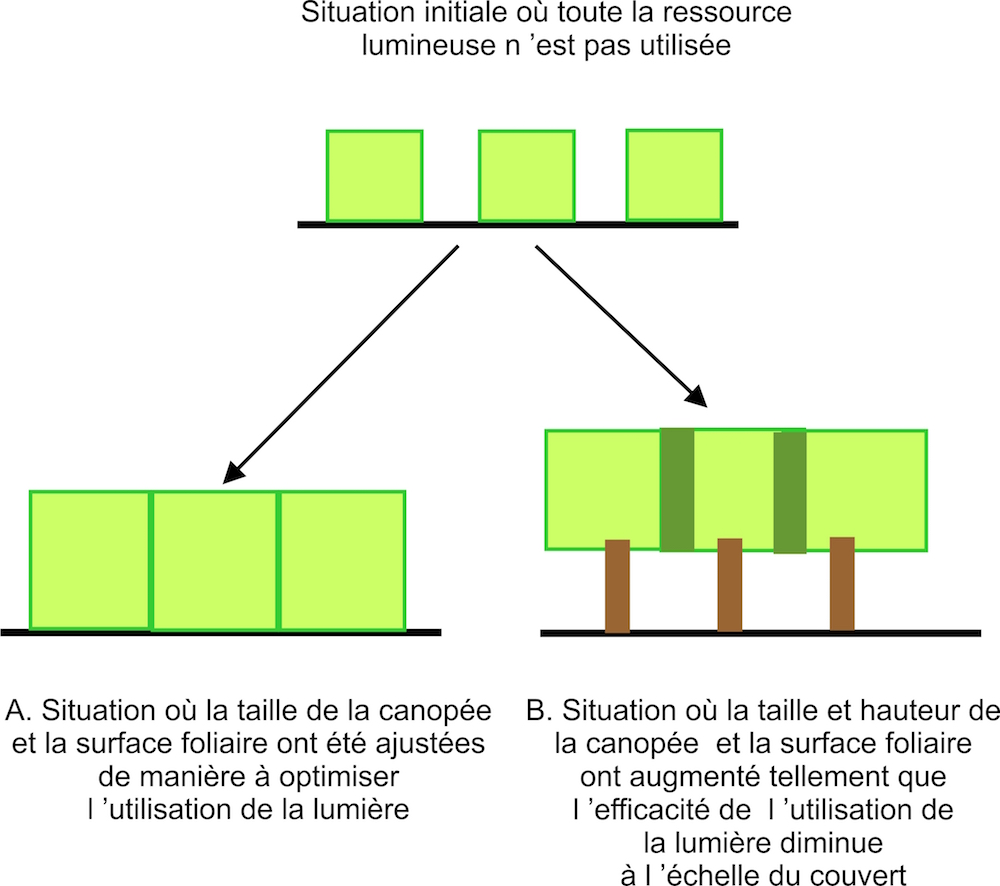
Figure 1
Figure 1 : Compétition pour la lumière au sein d’un couvert de plantes. Si la compétition et ses conséquences évolutives poussent à optimiser l’extension horizontale des canopées et la surface foliaire (cas A) la production de biomasse à l’échelle du couvert peut être optimisée. Si au contraire la compétition et l’évolution conduisent une extension horizontale et verticale de la canopée trop importante, une surface foliaire trop importante (cas B) la production de biomasse peut diminuer. Dans le cas B le vert sombre correspond à des zones où plusieurs plantes se font mutuellement de l’ombre et où la surface foliaire est plus élevée que la surface foliaire optimale. De même, dans le cas B l’apparition d’un tronc est aussi due à la compétition pour la lumière qui pousse évolutivement les plantes à aller chercher la lumière de plus en plus haut. La production d’un tronc pérenne requiert l’utilisation de ressource et conduit aussi à une perte d’efficacité.

Savane de Lamto en côte d’Ivoire.
Une strate arborée est en compétition avec une strate de graminées pour la lumière et les nutriments minéraux et l’eau du sol. © S. Barot

Champ de blé
Les pieds de blé sont en compétition pour la lumière et les ressources du sol. L’homme a sélectionné des variétés produisant la quantité optimale de racine et de surface foliaire pour optimiser la production de grains. © S. Barot
Il a été suggéré que, si la sélection naturelle tend à conduire à de telles situations non-optimales du point de vue de propriétés collectives comme la production primaire, la sélection artificielle* quant à elle – par exemple la sélection de variétés de plantes agricoles – peut permettre de maximiser la production agricole (Denison et al. 2003), c.-à-d. la biomasse de graines ou de fruits. Cela est possible car, dans ce cas, le critère de sélection d’une variété de plante est fondé sur la maximisation du rendement global d’un champ et non sur la maximisation du nombre de descendants viables d’une plante. On pense ainsi que les variétés cultivées ont petit à petit perdu la compétitivité qui avait été sélectionnée par la sélection naturelle chez les espèces sauvages dont elles sont les descendantes (Loeuille et al. 2013, Reynolds et al. 1994). Se faisant, les variétés modernes ne « gaspillent » plus de ressources en produisant plus de racines et de feuilles qu’il est nécessaire pour maximiser la production.
Application au système de recherche scientifique
Il faut bien sûr prendre beaucoup de précautions quand on cherche à appliquer aux sociétés humaines des résultats portant sur le fonctionnement des écosystèmes et des communautés d’organismes non-humains. Quels que soient les résultats de l’écologie et de l’écologie évolutive, les sociétés humaines peuvent en principe refuser de prendre des mesures maximisant l’efficacité de telle ou telle de leurs caractéristiques collectives (par exemple la production d’un pays) si ces mesures ne sont pas jugées acceptables pour des raisons morales ou philosophiques. L’espèce humaine peut en théorie décider de son chemin et n’a aucune raison d’imiter « bêtement » ce qui se passe dans la nature.
Cependant, l’impact écologique et évolutif de la compétition (présenté ci-dessus) montrent assez clairement que la compétition et la compétitivité ne sont pas une panacée pour améliorer et optimiser « l’efficacité des sociétés humaines » ou toute autre propriété émergeant des comportements humains individuels. Typiquement, la compétition est très souvent présentée comme un moteur naturel de la recherche et de la production de nouvelles connaissances. Ce leitmotiv a tellement été mis en avant dans la société en général (voir plus bas) et par la direction de la recherche que la communauté académique (y compris en écologie) a tendance à largement accepter cette idée.
On peut comprendre que le désir de faire de nouvelles « découvertes », d’être le premier à comprendre tel ou tel phénomène et d’acquérir une position académique (et sociale) plus élevée motive les chercheurs. Cela peut s’inscrire dans une émulation plutôt saine. Cependant, les résultats de l’écologie (voir plus haut) suggèrent que la seule compétition, si elle n’est pas contrebalancée par d’autres processus, a peu de chance d’optimiser la production de nouvelles connaissances (et l’utilisation de ces connaissances pour répondre aux besoins de la société) qui est une propriété émergente collective de l’ensemble des chercheurs et organismes de recherche.
En résumé, quand la compétition pousse à développer des théories sans cesse plus originales et plus en adéquation avec la réalité et à utiliser ces théories, la science et les sociétés humaines bénéficient de cette compétition. En revanche, quand la compétition pour des ressources limitantes nombreuses (financements, espaces de laboratoire, les meilleurs étudiants, accès aux médias…) conduit les chercheurs à dépenser toute leur énergie pour acquérir ces ressources et empêcher les autres chercheurs de les acquérir, il y a toutes les chances que la science et les sociétés humaines y perdent (moins de connaissances sont produites et les sociétés bénéficient moins de l’utilisation de ces connaissances).
Malheureusement, nous savons tous à quel point le système académique à l’échelle nationale et internationale est trop souvent devenu inefficace. Il peut paraître normal qu’un chercheur passe une partie de son temps à répondre à des appels d’offre compétitifs pour obtenir les financements pour des projets d’ampleur exceptionnelle. Il est moins normal que les chercheurs dépendent de tels appels d’offre pour effectuer leurs « recherches de base » au quotidien, ou que le taux de réussite de beaucoup d’appels d’offre soit inférieur à 10% (cas de l’ANR en 2014 et 2015, Huet 2016). Répondre à un appel d’offre, c’est aussi commencer à élaborer des nouvelles hypothèses et de nouvelles recherches. Cela peut donc être vu comme une activité normale du chercheur.

– – (© A. Teyssèdre )
Cela devient anormal quand le formalisme des appels d’offre (et en particulier leur volet administratif) devient tellement lourd que le temps nécessaire pour répondre à l’appel d’offre augmente exponentiellement et qu’on propose même aux chercheurs des formations pour répondre à tel ou tel type d’appel d’offre ou qu’on leur conseille de payer des entreprises privées pour faire une partie du travail. Dans tous ces cas, une partie grandissante des ressources des chercheurs, du temps de travail et, in fine, de l’argent public, est détournée de sa fonction première, faire avancer la recherche, afin de remplir des fonctions accessoires (cf. le regard n°15 de Jérôme Casas). Cela diminue l’efficacité collective du système de recherche, même si individuellement chaque chercheur n’a pas le choix et doit investir plus de temps qu’il ne devrait être nécessaire dans des tâches ne produisant pas directement de nouvelles connaissances : pour trouver des ressources, pour maintenir sa compétitivité… C’est exactement ce qui se passe quand la sélection naturelle sélectionne chez une plante une production de surface foliaire améliorant potentiellement sa production individuelle de biomasse et sa propre compétitivité mais diminuant la production collective d’une population d’individus ayant tous la même stratégie.

– –
Les excès du système de publications sont aussi largement reconnus. L’ensemble du système académique est polarisé par le nombre d’articles publiés (par chercheurs, équipes, laboratoires ou institutions de recherche) et le nombre de citations de ces articles. Ainsi, le recrutement des chercheurs, l’évaluation des laboratoires et des projets soumis aux appels d’offre dépendent très étroitement du nombre de publications publiées par les individus. Cela engendre une compétition très intense pour une « ressource limitante », la place dans les journaux scientifiques et les citations.
Bien sûr, il est indispensable que les chercheurs publient le résultat de leurs recherches, mais cette compétition excessive a de nombreux effets négatifs : (1) Les chercheurs tendent à publier leurs résultats dans des publications de plus en plus courtes et nombreuses, sans nécessairement se préoccuper de la qualité de ces résultats. (2) Beaucoup de publications sont redondantes. (3) l’abondance des publications fait qu’un chercheur ne peut plus lire toutes les publications de son domaine. (4) Cette compétition pour les publications pousse de nombreux chercheurs à tricher en publiant des résultats faux, en inventant des données ou en sur-interprétant leurs résultats.
Enfin, la recherche se base largement autant sur la collaboration que sur la compétition. Au minimum, tout projet de recherche demande de très nombreuses compétences qu’aucun chercheur ne peut posséder à lui seul. D’une manière plus générale, la recherche est basée sur l’échange d’idées et de points de vue qui seul permet la progression des connaissances par amélioration progressive des théories. De plus, l’écologie scientifique a de plus en plus besoin de faire des synthèses, des méta-analyses, des projets de grande ampleur basés sur de très grosses bases de données. C’est la seule manière d’aboutir à des théories plus générales, de devenir capable de faire des prédictions plus précises. De nouveau, ces nouvelles formes de recherche demandent des collaborations plus larges et une meilleure coordination des activités des chercheurs conduites dans un but commun (standardisation des protocoles, très grosses expériences, observatoires, mise en réseau, construction de bases de données communes…). Cela montre bien que la seule compétition ne peut pas servir de moteur universel à la recherche.
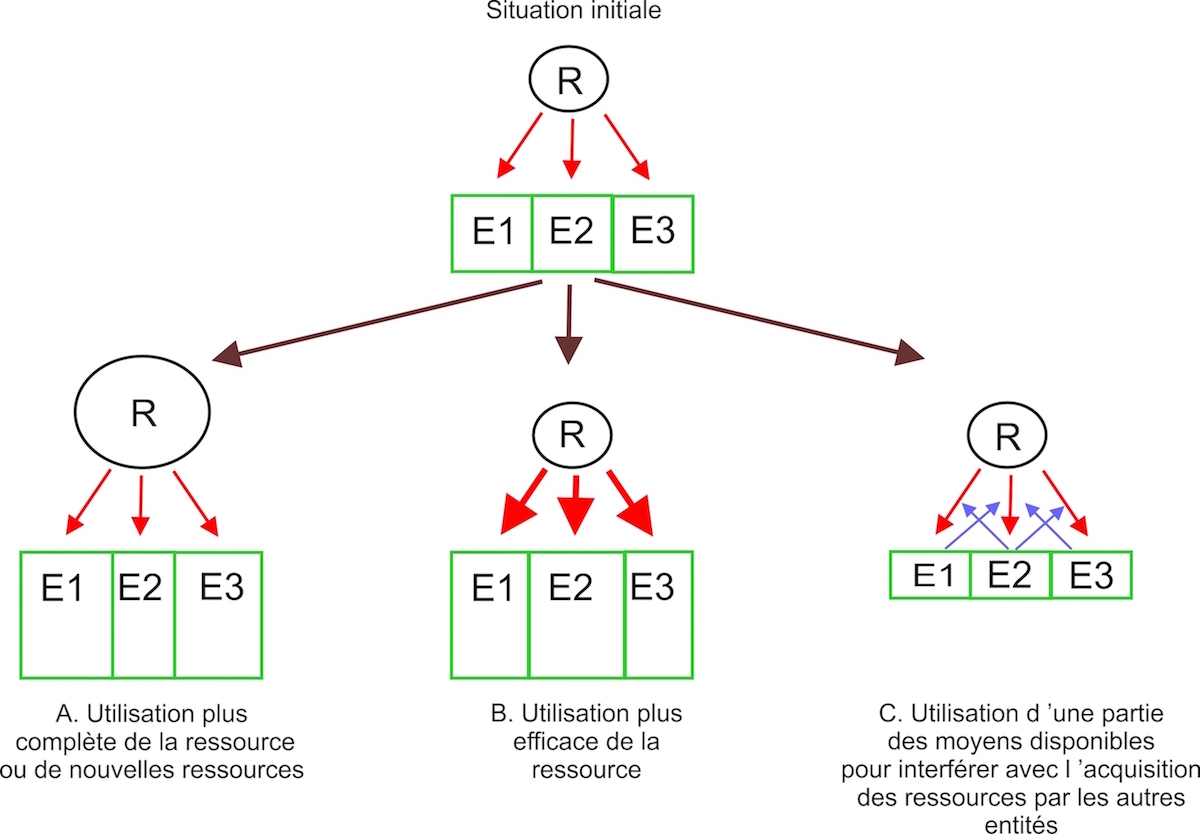
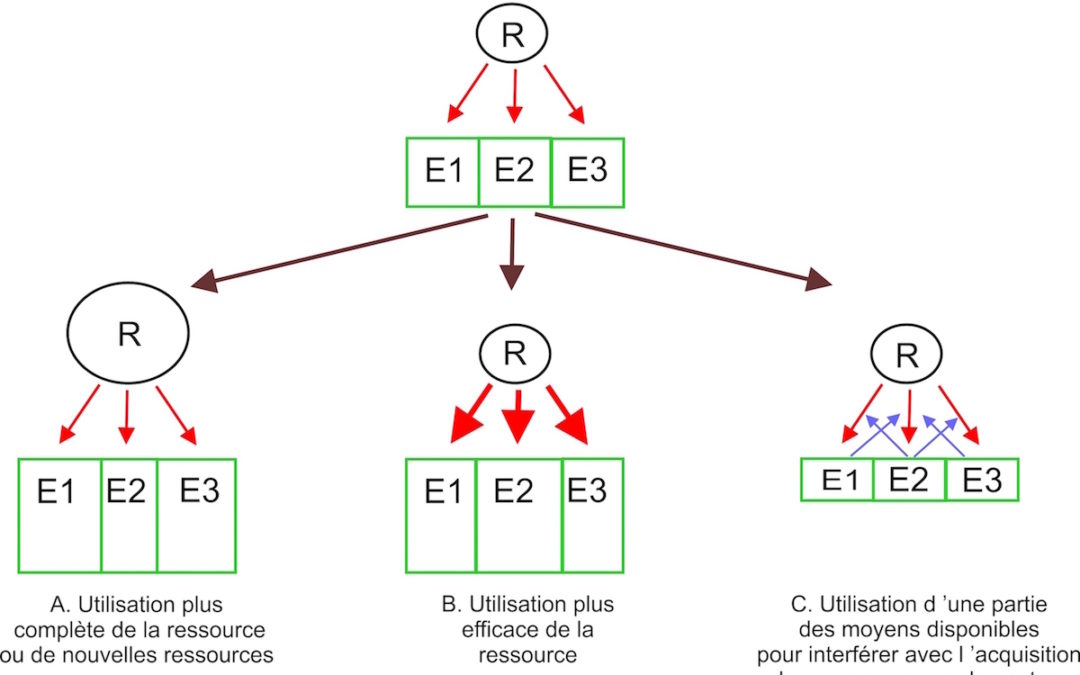
Figure 2
Figure 2: Effet de la compétition pour une ressource (R) entre 3 entités (E1, E2, E3) sur la taille de ces entités et leur capacité de production (taille des rectangles verts). La compétition peut pousser les entités à modifier leur stratégie de façon à ce qu’elles augmentent leur accès à la ressource. Le cas A (la compétition conduit à une utilisation plus complète de la ressource) et le cas B (la compétition conduit à une plus grande efficacité de l’utilisation de la ressource) conduisent à une plus grande capacité collective de production des entités. Dans le cas C (la compétition pousse les entités à interférer avec la capacité d’acquisition de la ressource des autres entités) cela conduit au contraire à une diminution de la capacité collective de production des entités. Ce schéma peut représenter la compétition entre organismes vivants pour une ressource qui est transformée en biomasse. Il peut aussi représenter la compétition entre chercheurs (ou entités de recherche) qui transforment une ressource financière en nouvelles connaissances. Dans ce cas, le système de financement peut parfois conduire à une diminution de l’efficacité de l’utilisation de la ressource: c’est l’opposé du cas B. Certains aspects du fonctionnement académique peuvent ainsi correspondre au cas C. Enfin, le schéma peut correspondre à la compétition entre plusieurs entreprises pour un marché. Dans ce cas, la taille des rectangles verts symbolise les services fournis (en prenant en compte la quantité, la qualité, les prix) par l’entreprise et la ressource est l’ensemble des clients/consommateurs. Si la compétition conduit à la production de nouveaux produits ou une production plus efficace d’un produit, on est dans le cas A ou B. Si la compétition se traduit par l’utilisation d’une partie des moyens de l’entreprise pour interférer avec les autres entreprises sans jouer sur la production, cas de la publicité, on est dans le cas C. Bien sûr, les mécanismes conduisant un organisme à changer de stratégie (sélection naturelle), un chercheur à essayer d’obtenir plus de financement (désir de développer des recherches et de se maintenir dans le système académique malgré la compétition) et une entreprise à gagner des parts de marché (désir des acteurs d’augmenter leurs revenus) sont très différents.
Application aux sociétés humaines
Si on peut essayer d’appliquer les résultats de l’écologie sur la compétition au système de recherche, il est aussi tentant de les utiliser pour interpréter le fonctionnement de l’ensemble de notre société. Comme dans la recherche, la société, du moins notre société moderne occidentale, a tendance à survaloriser la compétition et la compétitivité et, surtout, à mettre en avant le fait que la compétition est la meilleure solution pour augmenter « l’efficacité de notre société ». De nouveau, les résultats des recherches en écologie suggèrent que cela n’est pas le cas.
Les conséquences de la compétition à l’échelle de la société devraient au moins dépendre des mécanismes impliqués. Comme pour les plantes, si la compétition conduit les individus/ les entreprises/ les pays à améliorer l’usage des ressources (par exemple en produisant plus de produits manufacturés à un moindre coût) la compétition peut bénéficier à l’ensemble de la société. Si, au contraire, la compétition pousse les individus/ les entreprises/ les pays à utiliser une partie des ressources (financières, énergétiques…) pour augmenter leur compétitivité sans que cela se traduise par une amélioration (qualitative ou quantitative) de la production, cela a peu de chance de bénéficier à la société dans son ensemble.

Times Square, New York
© CC BY-SA-1.0
Ainsi, on a pu estimer que la publicité correspond souvent à plus de 10% du prix de vente de certains produits et parfois même jusqu’à 2/3 de ce prix (Wikipédia). La publicité représente bien un coût pour le consommateur. Probablement, selon son créneau économique, une entreprise peut ne pas avoir le choix. Pour survivre, elle peut avoir besoin d’investir dans la publicité, parce que les entreprises avec lesquelles elle est en compétition font de la publicité. Il s’agit donc de nouveau d’une course aux armements, comme dans le cas du chercheur qui doit répondre à de très nombreux appels d’offre, comme dans le cas d’une plante qui investit trop dans sa surface foliaire « pour » faire de l’ombre à ses voisins. Le coût total de la publicité est énorme. On estime ainsi la valeur totale payée en France par les annonceurs de publicité à environ 10 milliards d’euros par an (environ 20 fois le budget d’intervention de l’ANR qui était d’environ 0,5 Milliards d’euros en 2014 ; plus que le budget consacré en France à la recherche hors enseignement supérieur, environ 8 milliards d’euros).
De nouveau, une entreprise peut bénéficier de la publicité si celle-ci lui permet de gagner des parts de marché, i.e. de gagner la compétition. Mais la société dans son ensemble ne tire pas de bénéfice du fait que telle ou telle entreprise gagne des parts de marché grâce à la publicité.
Enfin, nous avons tous l’habitude de bénéficier de « services gratuits » financés par la publicité. C’est le cas par exemple des journaux d’information « gratuits » et de nombreux services internet. Mais bien sûr ces services ne peuvent pas être « gratuits »; there’s no such thing as a free lunch (titre d’un livre de Milton Friedman et slogan des tenants du libéralisme économique, Friedman 1975). Les annonceurs payent pour que leurs publicités soient diffusées dans les journaux et sur internet et ce coût doit être répercuté sur le prix des biens et des services qu’ils produisent. On peut aussi penser que la publicité commerciale rend un service en informant les consommateurs. Mais il est alors facile d’argumenter qu’il ne s’agit pas d’une information non-biaisée. On peut même argumenter qu’il s’agit souvent de désinformation. Si je veux choisir en toute connaissance de cause un produit, mon choix sera probablement mieux fondé s’il s’inspire des articles d’un journal de consommateurs (comme « 60 millions de consommateurs » ou « Que choisir ?») que sur des publicités. Bien sûr, on peut développer beaucoup d’autres arguments contre la publicité (coût environnemental, argument moraux…), mais il me parait pertinent de développer ici cet argument basé sur son coût économique.
Conclusion
Je pense qu’il est important que la science écologique fasse bénéficier la société de toutes ses avancées de toutes les manières possibles. Le but de ce texte est bien d’aller dans ce sens. L’écologie scientifique a permis d’atteindre une compréhension importante du fonctionnement et de la dynamique de systèmes écologiques très complexes. Elle a ainsi développé une approche systémique inégalée, qui pourrait être transposée pour étudier et interpréter de nombreux aspects du fonctionnement des sociétés humaines, et en particulier l’économie. Par exemple la théorie des jeux est déjà à la fois utilisée en écologie, en écologie évolutive et en économie (voir par exemple Rabin 1993).
Enfin, l’écologie étudie aussi les interactions positives entre organisme, comme le mutualisme, et ces travaux montrent que ce type de relation joue un rôle crucial pour de nombreux organismes et les écosystèmes qui les hébergent. Par exemple, les producteurs primaires dépendent fortement de symbioses* avec des champignons pour leur croissance et leur survie (Selosse and Le Tacon 1998). Les mycorhizes* forment ainsi des réseaux explorant très finement l’ensemble du sol et peuvent ainsi absorber les nutriments minéraux avec une très grande efficacité. Les mycorhizes étant reliées aux racines des plantes elles permettent aux plantes d’acquérir plus de ressources (des nutriments minéraux). Les plantes sont des producteurs primaires et peuvent en échange leur fournir de la matière organique. Les symbioses mycorhiziennes sont ainsi très importantes pour les deux partenaires et elles jouent un rôle crucial pour la production primaire terrestre.
L’écologie a aussi étudié les conditions permettant à des symbioses de s’établir et de se maintenir (Denison 2000, de Mazancourt and Schwartz 2010) ou permettant de diminuer l’intensité de la compétition (Bachmann et al. 2013, Barot et al. 2016). Même si la description du mutualisme va au-delà des objectifs de cet article, il est certain que l’étude de ces interactions positives, et pas seulement celle des interactions négatives comme la compétition, pourrait aussi éclairer le fonctionnement des sociétés humaines, et éventuellement suggérer des pistes de solutions à différents problèmes économiques et sociaux. Comme suggéré plus haut, la coopération est clairement un des moteurs de la recherche. La coopération est d’une manière générale essentielle au bon fonctionnement des sociétés humaines.
Glossaire
Mycorhizes : les mycorhizes sont des champignons en symbioses avec les plantes. Les mycorhizes, liées aux racines, fournissent aux plantes des nutriments minéraux et les plantes fournissent en retour de la matière organique.
Niche écologique : ensembles des conditions environnementales (biotiques et abiotiques) permettant à une espèce de vivre et de se reproduire.
Population : ensemble d’individus de la même espèce en interaction dans une même zone
Production primaire : quantité de biomasse produit par les producteurs primaires d’un écosystème, par exemple les plantes.
Ressource abiotique : ressource n’ayant pas une origine biologique utilisée par un organisme pour sa croissance. Ainsi l’eau et les nutriments minéraux constituent des ressources abiotiques pour les plantes.
Sélection artificielle : processus permettant aux humains de sélectionner chez des plantes ou des animaux domestiqués des caractéristiques particulières (par exemple une production élevée de graines ou de lait). Contrairement à la sélection naturelle, il est possible de sélectionner n’importe quelle caractéristique, y compris des caractéristiques collectives comme la production de biomasse, même si cette caractéristique n’est pas liée au nombre de descendants.
Sélection naturelle : processus à la base de l’évolution darwinienne par lequel les individus les mieux adaptés aux conditions environnementales ont plus de descendants, ce qui tend à permettre aux caractéristiques de ces individus, i.e. des adaptations, de se répandre dans leur population.
Symbiose : interaction positive, durable et intime entre deux organismes qui tirent réciproquement bénéfice de cette interaction.
Bibliographie
Bachmann, H., M. Fischlechner, I. Rabbers, N. Barfa, F. B. dos Santos, D. Molenaar, and B. Teusink. 2013. Availability of public goods shapes the evolution of competing metabolic strategies. Proceedings of the National Academy of Sciences of the USA 442:75-78.
Barot, S., S. Bornhofen, S. Boudsocq, X. Raynaud, and N. Loeuille. 2016. Evolution of nutrient acquisition: when space matters. Functional Ecology 30:283-294.
Boudsocq, S., S. Barot, and N. Loeuille. 2011. Evolution of nutrient acquisition: when adaptation fills the gap between contrasting ecological theories. Proc. Royal. Soc London B 278:449-457.
de Mazancourt, C. and M. W. Schwartz. 2010. A resource ratio theory of cooperation. Ecology Letters 13:349-359.
Denison, R. F. 2000. Legume sanctions and the evolution of symbiotic cooperation by rhizobia. American Naturalist 136:567-576.
Denison, R. F., J. M. Fedders, and B. L. Harter. 2010. Individual fitness versus whole-crop photosynthesis:solar tracking tradeoffs in alfalfa. Evolutionary Applications 3:466-472.
Denison, R. F., E. T. Kiers, and S. A. West. 2003. Darwinian agriculture: when can humans find solution beyond the research of natural selection? The quaterly review of biology 78:145-168.
Friedman, M. 1975. There’s no such thing as a free lunch. Open Court Publishing Company.
Gersani, M., J. S. Brown, E. E. O’Brien, G. M. Maina, and Z. Abramsky. 2001. Tragedy of the commons as a result of root competition. Journal of Ecology 89:660-669.
Hardin, G. 1960. The competitive exclusion principle. Science 131:1292-1297.
Hardin, G. 1968. Tragedy of commons. Science 162:1243-&.
Huet, S. 2016. Politique de la recherche : démission de masse à l’ANR. Blog {Science2}, Le Monde.
Loeuille, N., S. Barot, E. Georgelin, G. Kylafis, and C. Lavigne. 2013. Eco-evolutionary dynamics of agricultural networks: implications for a sustainable management. Advances in Ecological Research 49:339-435.
Malthus, T. R. 1798. An essay on the principle of population. J. Johnson, London.
Parvinen, K. 2005. Evolutionary suicide. Acta Biotheoretica 53:241-264.
Rabin, M. 1993. Incorporating Fairness into Game Theory and Economics. The American Economic Review 83:1281-1302.
Reynolds, M. P., E. Acevedo, K. D. Sayre, and R. A. Fischer. 1994. Yield potential in modern wheat varieties: its association with a less competitive ideotype. Field Crops Research 37:149-160.
Selosse, M.-A. and F. Le Tacon. 1998. The land flora: a phototroph-fungus partnership? Trends in Ecology & Evolution 13:15-20.
Stearns, S. C. 1992. Quantitative genetics and reaction norms. Pages 40-71 The evolution of life histories. Oxford University Press, Oxford.
Tilman, D. 1982. Resource competition and community structure. Princeton University Press, Princeton, New Jersey.
Turchin, P. 2001. Does population ecology have general laws? Oikos 94:17-26.
—-
‘Regard’ édité par Anne-Violette Lavoir, illustré et mis en ligne par Anne Teyssèdre
Sébastien, ton article est vraiment très intéressant. Paradoxalement, il s’applique dans le cadre des systèmes compétitifs de la recherche la plus poussée et la plus actuelle mais comme tu le suggères il s’applique aussi dans les systèmes les plus conservateurs, favorisant à terme une « extinction » ou « suicide ».
L’académisme et le système universitaire et scientifique français (mais ils ne sont pas les seuls dans ce cas) sont dominés par des mandarins qui détiennent le pouvoir de bloquer les promotions, les capacités d’innovation parce que la compétition, c’est eux qui l’organisent à leur profit : recrutements contrôlés pour leurs féodaux, appels d’offre truqués dans les comités scientifique où les membres se servent eux-mêmes en s’attribuant à eux-mêmes ou à leurs équipes les financements etc.
Bref, une fausse compétition qui ne dit pas son nom et qui se transforme, comme dans les écosystèmes qui perdent en biomasse, par une domination de quelques clans souvent incapables de rénover la science.
Merci pour cet article très revigorant puisque très lucide et qui renouvelle en quoi une science humaine lucide doit s’éclairer à constater quels sont les processus et les dynamiques à l’oeuvre en effectuant un travail critique réflexif… y compris sur les comportements des disciplines scientifiques elles-mêmes. Les travaux des éthologues viennent aussi nous aider à comprendre les comportements sociaux… y compris des systèmes scientifiques.
Bonjour,
Merci pour cet excellent article qui a le mérite d’englober la société humaine dans le processus auquel elle appartient et dans l’espace vivant de l’écosystème. Je pense qu’implicitement nos sociétés occidentales ont oublié qu’elles font partie d’un ensemble d’espèces et que la séléction est à l’oeuvre, et que l’action des individus à des répercussions à toutes les échelles. Ce qui est paradoxal c’est que l’homme se voit comme maître de sa destinée, comme hors-nature, mais dès lors qu’il lui est demandé de réflechir, de choisir, de se limiter, alors la force de l’habitude et l’inertie lui font dire que « non, ce n’est pas possible ».
Nous sommes plein de paradoxes (la publicité en est un je trouve) et peut-être que ce que nous cherchons depuis toujours comme étant la « marque de l’homme » pourrait être cette capacité à choisir la résilience collective plutôt que l’épuisement pour motif individuel..
Encore merci !
Bien cordialement
Merci pour ces commentaires positifs, Pierre et Abludivine.
J’avais peur que le texte ne soit pas assez clair, mais apparemment cela n’est pas le cas.