
La Société Française d’Ecologie et d’Evolution (SFE2) vous propose ce regard de Patrick Giraudoux et Jean-Dominique Lebreton, respectivement Professeur à l’Université de Bourgogne Franche-Comté et Directeur de recherche au CNRS, sur les notions d’équilibre et de santé des écosystèmes.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
———-
La notion de santé des écosystèmes, ou
‘Zombies écologiques : comment des idées mortes
vivent encore parmi nous’
par Patrick Giraudoux(1) et Jean-Dominique Lebreton(2)
(1) : Membre honoraire de l’Institut Universitaire de France, Professeur d’écologie, Laboratoire Chrono-environnement, Université de Bourgogne Franche-Comté et CNRS,
https://chrono-environnement.univ-fcomte.fr/acteurs/annuaire/article/giraudoux-patrick
(2) : Membre de l’Académie des sciences, Directeur de recherche CNRS, CEFE, UMR 5175 CNRS-Université de Montpellier-Université Paul Valéry.
.
Ce regard est une révision actualisée par P. Giraudoux et J.D. Lebreton, avec l’appui éditorial d’A. Teyssèdre et S. Barot, d’un chapitre d’ouvrage paru en 2014 (Giraudoux, 2014). Le titre est un clin d’œil à l’ouvrage de J. Quigging ‘Zombie economics: how dead ideas still walk among us.’ Princeton University Press, Princeton, 2010.
——-
Mots clés : Biodiversité, changement, conservation, dynamique, équilibre, évolution, paysage, santé, socio-écosystème, système multi-potentiel, service écosystémique.
——–
- Équilibre et santé des écosystèmes : deux concepts ambigus
- Le regard de Costanza
- De la durée des écosystèmes
- Santé des écosystèmes et services écosystémiques
- Des écosystèmes, du vivant et de leur évolution
- Des zombies écologiques
- Conclusion
- Bibliographie
- Regards connexes
- Forum de discussion sur ce regard
——-
Équilibre et santé des écosystèmes : deux concepts ambigus
La notion de « santé des écosystèmes », popularisée par les médias, date du début du XIXème siècle. On en retrouve trace dans le célèbre « Almanach d’un Comté des Sables » d’Aldo Leopold un des ouvrages fondateurs de l’éthique environnementale, de la pensée écologiste, et de la biologie de la conservation (Leopold, 1949). Elle est encore souvent utilisée par la communauté des chercheurs en écologie : ainsi un journal scientifique s’appelle « Ecosystem Health and Sustainability » (https://www.tandfonline.com/loi/tehs20).
Un état de « bonne santé » est, à l’extrême, parfois considéré comme un état d’équilibre souhaitable et permanent sur lequel l’homme n’interviendrait pas, ou seulement pour le perturber (Zimmerman & Cuddington, 2007). Cet état d’équilibre idéalisé et cette métaphore de santé des écosystèmes ne résistent pas à l’analyse critique. Cette vision idéalisée d’une nature dont Homo sapiens serait le seul perturbateur, et qui serait capable de revenir spontanément à un équilibre unique et idéal, est en effet contredite de multiples manières : les systèmes écologiques, avec ou sans l’espèce humaine, sont tous dans un état provisoire qui est le résultat de dynamiques complexes, sujettes à de nombreuses interactions, dès lors qu’une durée suffisante est considérée. Ces dynamiques sont suffisamment complexes pour rendre toute prédiction à long terme sur l’état de ces systèmes écologiques difficile, pour ne pas dire impossible, sauf en ce qui concerne un certain nombre d’invariants fonctionnels structurels (voir Simberloff et Wilson, 1969, pour un exemple historique d’études sur ce sujet).
De nombreux états durables sont possibles pour un assemblage donné d’espèces, et des fonctions écologiques identiques peuvent être réalisées avec des assemblages d’espèces différentes (Naeem, 1998). Les avantages et désavantages des différents états de systèmes écologiques sont essentiellement perçus d’un point de vue anthropocentré. Dans de tels anthropo-écosystèmes, et dans un contexte biogéographique donné, la « santé » des écosystèmes et leur équilibre dynamique et provisoire apparaissent principalement comme un choix de combinaisons de contraintes économiques et un consensus social, parmi une grande variété d’équilibres dynamiques possibles que les composantes non-humaines de l’écosystème pourraient installer « durablement » à un temps donné. Un défi important pour la recherche est certainement d’apporter les concepts scientifiques nécessaires à la gestion de tels systèmes au sein desquels de multiples interactions se combinent pour rendre toute gestion statique ou fixiste inappropriée, y compris quand elle concerne la biodiversité.

.
Figure 1 : Retrait forestier au sud-ouest de Pontarlier, vu du même point d’observation.
a : au XIXème siècle (peinture anonyme réalisée entre 1838 et 1865, collection Musée municipal de Pontarlier) ;
b : en août 2013.
Le regard de Costanza
Costanza et ses coauteurs (1992), dans un ouvrage pluridisciplinaire associant écologues, philosophes et économistes, ont été parmi les premiers et les rares à tenter une analyse objective du concept de santé des écosystèmes (concept décliné, pour un certain nombre de composantes des écosystèmes, dans le récent regard RO6 de M. Duru). Selon Costanza, la santé d’un écosystème devrait être jugée selon trois propriétés indépendantes : sa vigueur (c’est-à-dire une mesure de l’activité de l’écosystème, de son « métabolisme primaire » -sic- ou de sa productivité primaire), son organisation (c’est-à-dire le nombre et la diversité des interactions entre ses composantes) et sa résilience (c’est-à-dire sa capacité à retrouver sa structure et ses fonctions après un stress) (Costanza, 2012). La santé d’un écosystème serait donc la combinaison de ces trois propriétés.
Costanza et al. (1992) précisent qu’un « système écologique est en bonne santé, et libre de “syndrome de détresse” s’il est stable et durable, c’est à dire s’il est actif en maintenant son organisation et son autonomie au cours du temps, et est résiliant au stress » (notre traduction). Costanza (2012) ajoute qu’un tel écosystème est qualifié de « durable s’il survit sur un temps spécifié (non infini) » et est en « bonne santé » s’il « procure une gamme de services écosystémiques ». Cette définition de la santé d’un écosystème se heurte à deux difficultés. La première est qu’elle implique, au-delà du grain minimal relativement court des étapes d’une succession écologique classique, dont les durées sont variables selon le type d’écosystème, une durabilité sur un temps inévitablement arbitraire. La deuxième est qu’elle oblige à caractériser l’écosystème par les services écosystémiques qu’il procure, classés par définition de manière anthropocentrée, comme biens communs, vitaux ou utiles pour l’être humain, les autres espèces et les activités économiques (Millennium Ecosystem Assessment, 2005).
De la durée des écosystèmes
On estime que les formes vivantes actuelles ne représentent que 4 à 6% des espèces ayant existé depuis l’apparition de la vie il y a 3,8 milliards d’années (Mace et al., 2005). Un grand nombre d’écosystèmes se sont bien sûr succédés depuis, dont les couches géologiques gardent trace, perturbés par des accidents dont témoignent cinq crises majeures d’extinction d’espèces depuis le Cambrien. La crise d’extinction la plus récente, datée de 65 Millions d’années, à la transition Crétacé-Tertiaire est marquée notamment par la disparition des grands dinosaures (dinosaures non aviens). Elle est généralement attribuée à la chute d’une météorite sur le Yucatan qui aurait modifié de façon brutale et considérable le climat terrestre. A ces crises s’ajoute celle que nous vivons actuellement et dont notre espèce est responsable (e.g. Ceballos et al., 2015 ; Teyssèdre, 2004).
Chaque écosystème, sur une échelle de temps longue, a inévitablement été remplacé par un écosystème différent comportant d’autres espèces et des interactions et mécanismes de régulation réarrangés. Quand de grands groupes tels que les mammifères succèdent en quelques centaines de milliers d’années à de grands groupes comme les dinosaures, cette affirmation d’instabilité des écosystèmes n’étonne personne. Les changements, cependant, peuvent être beaucoup plus rapides. Ainsi, plus récemment, les formations forestières postglaciaires ont été remplacés progressivement par des paysages agricoles, par déboisement successif des terres les plus favorables à l’agriculture et à l’élevage, optimisant sous contrainte du relief et de la géologie les ressources disponibles, et créant des assemblages d’espèces et des interactions constituant de facto de nouveaux écosystèmes.
Tous les paysages actuels souvent qualifiés de « naturels » parce que verts ou inhabités, sont en fait le résultat d’un compromis entre exploitation des ressources locales par l’homme et potentiel biologique local. Le « naturel » n’est ici qualifié ainsi que par opposition à l’urbain et au « bétonné », et devrait plus judicieusement être qualifié de rural, et tout au plus de « vert » sur un gradient qui va du gris du béton au vert de la végétation.
Encore ne faut-il pas trop s’intéresser aux archives même récentes, pour y voir une stabilité. La peinture réaliste, outre ses vertus propres, possède celle de nous offrir une vision brute de la vie quotidienne et des paysages dès le XIXème siècle. Une simple visite du musée de Pontarlier ou du musée Courbet à Ornans, par exemple, convaincra le lecteur des profondes transformations de l’occupation biologique de cet espace rural sur moins d’une centaine d’années (Fig. 1). Dans une région où, de mémoire de franc-comtois, la race montbéliarde et le comté sont les piliers éternels et homogènes de l’agriculture de montagne, on est étonné de constater que dans les tableaux du milieu du XIXème siècle de Courbet et d’Antoine Beau, son maître, ni les paysages de montagne aux flancs déboisés en faveur des (sur)pâturages, ni les bovins quelque peu faméliques, ou les porcins bicolores, de diverses races surprenantes (voir, entre autres le tableau « Retour des paysans de la foire de Flagey », Fig. 2 ci-dessous) ne ressemblent à ceux qui caractérisent la Franche-Comté dans l’actuelle mémoire collective.

.
Figure 2 : Paysans de Flagey revenant de la foire (1850 – 1855), Gustave Courbet. Musée des beaux-arts et d’archéologie de Besançon.
Peut-on dire, à chacune des étapes écologiques schématiquement décrites plus haut, que les écosystèmes étaient en meilleure santé, sous critère de vigueur, d’organisation et de résilience ? Rien ne le prouve et le fait même que ces écosystèmes aient pu évoluer jusqu’à maintenant, préparant la vigueur, l’organisation et la résilience des écosystèmes du temps présent, prouve assez directement qu’ils ont assuré, au cours du temps, à travers le changement une continuité biologique fonctionnelle.
Santé des écosystèmes et services écosystémiques
“A healthy system should sustainably provide a range of ecosystem services” (Costanza, 2012). Ces services écosystémiques recouvrent une gamme de “services” assez large, classés en 4 grands types (Millenium Ecosystem Assessment, 2005, et voir le regard n°4) : i) approvisionnement humain (eau, nourriture, bois, fibre, etc.), ii) régulation (inondation, tempêtes, maladies, ravageurs et parasites, qualité de l’air, etc.), iii) culturels (esthétique, récréation, etc.), iv) soutien (ensemble des processus écologiques nécessaires au maintien des autres services).
La notion de service écosystémique est donc par définition totalement anthropocentrée. Un écosystème est en bonne santé s’il produit un nombre (lequel ?) de « services » assumés utiles pour les humains, sans que le nombre d’êtres humains soit pris en compte : les services attendus peuvent-ils être les mêmes quel que soit ce nombre, et tout type de niveau de vie ?
Par ailleurs, l’évolution darwinienne nous semble peu prise en compte par cette approche (mais voir Faith et al., 2010 ; Sarrazin et Lecomte, 2016). Non que les recherches sur les services écosystémiques et la santé des écosystèmes nient l’existence de l’évolution, mais elles se placent, dans la pratique, dans un contexte de court ou moyen terme où l’évolution darwinienne agit moins vite que les dynamiques des écosystèmes sous l’effet de la pression anthropique. Or, l’évolution du vivant n’est-elle pas centrale dès lors que les questions de durée et de durabilité sont considérées prioritairement ? En d’autres termes, peut-on penser la durée sans penser les processus qui prévalent dans cette durée ? Nous avons vu plus haut que les écosystèmes avaient une durée de vie limitée. Nous avons vu également que c’était le cas des espèces qui les composent.
Des écosystèmes, du vivant et de leur évolution
Objet mental idéal, l’espèce est classiquement définie comme « une population ou un ensemble de populations dont les individus peuvent effectivement ou potentiellement se reproduire entre eux et engendrer une descendance viable et féconde, dans des conditions naturelles » (Mayr, 1942). A l’intérieur même d’une espèce, la structure génétique d’une population est loin d’être homogène, et de nouveaux allèles apparaissent sans cesse par le jeu des mutations. On mesure de mieux en mieux la complexité, l’hétérogénéité et la plasticité des génomes et de leur expression. A cette diversité génétique correspond une diversité de formes et de fonctions, et de là une diversité d’interactions (parasitisme, mutualisme, compétition, prédation). Ces différentes diversités qui constituent la « biodiversité » sont loin d’être inventoriées. Chaque écosystème est bâti avec ces briques : chacun des éléments et chaque écosystème évolue donc sans cesse sous la pression de l’hétérogénéité des conditions physiques dans le temps et dans l’espace. La définition même du vivant et de la biodiversité s’oppose donc à toute vision fixiste des écosystèmes.
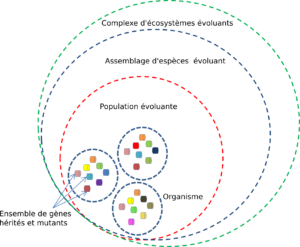
.
Figure 3 : Mobilis in mobile : emboîtement des niveaux d’organisation du vivant (des interactions sont spécifiques à chaque niveau) évoluant en permanence.
Des zombies écologiques
Si donc l’on s’accorde sur le constat de l’instabilité des écosystèmes à des échelles de temps et d’espace variées (Fig. 3), que reste-t-il de la notion d’équilibre écologique et de santé des écosystèmes, et à quelle(s) échelle(s) doivent s’appliquer d’éventuels principes de gestion ? En d’autres termes, ce qu’on appelle « environmental stewardship » (Chapin et al., 2009), c’est-à-dire le devoir qu’aurait l’humanité de gérer durablement les écosystèmes dont elle hérite, dont elle fait partie, et qu’elle lègue aux générations futures, est-il compatible avec une vision apparemment statique voire fixiste des écosystèmes et des espèces, supposés invariants dans la pratique (mais pas dans la théorie)? Comment échapper à ce dilemme qui, pour préserver le capital évolutif que représente la biodiversité (aux échelles variées où elle peut être mesurée) et stopper son érosion, nous conduit à chercher à la figer en l’état, tout en étant certain que la biodiversité du futur n’est pas la biodiversité du présent, et que les processus dynamiques qui sont responsables de cette biodiversité font que nous ne nous baignons jamais deux fois dans la même rivière ?
Dans une hiérarchie d’organisation, la dynamique des écosystèmes à une échelle spatiale donnée dépend donc en bonne part de celles mesurées à une échelle plus large et détermine celles mesurées aux échelles plus locales. Quel principe pratique de gestion de la biodiversité en tirer ? Un écosystème, nécessairement à une échelle d’espace et de temps donnée explicitement, serait donc en « bonne santé » s’il préserve les dynamiques, les capacités évolutives et les services écosystémiques des systèmes qui l’incluent et de ceux qu’il inclut. Dans un tel contexte fluide, et avec encore peu de concepts éprouvés et consensuels pour servir un projet encore très flou d’environmental stewardship, le chercheur, et à plus forte raison le gestionnaire, sont assez désarmés.
Veiller à la santé des écosystèmes supposerait donc, au-delà d’une préservation à court terme, de veiller à préserver leurs capacités évolutives en accompagnant leurs transitions (Fig. 4) ? Dans quelle direction ? On peut adopter une position anthropocentrée : la ou les directions qui maintiennent la plus vaste gamme de services écosystémiques et de possibilités d’évolution. Ce qui revient à préserver le futur à la seule lumière du présent. Peut-on être moins anthropocentré ? Difficilement.
Les notions d’équilibre écologique et de santé des écosystèmes, indéfinissables objectivement, sont donc sinon des concepts morts-vivants (des zombies), bien plutôt révélatrices de consensus sociaux qui s’établissent à un moment donné sur la gamme de services durables qu’on attend des dits systèmes.
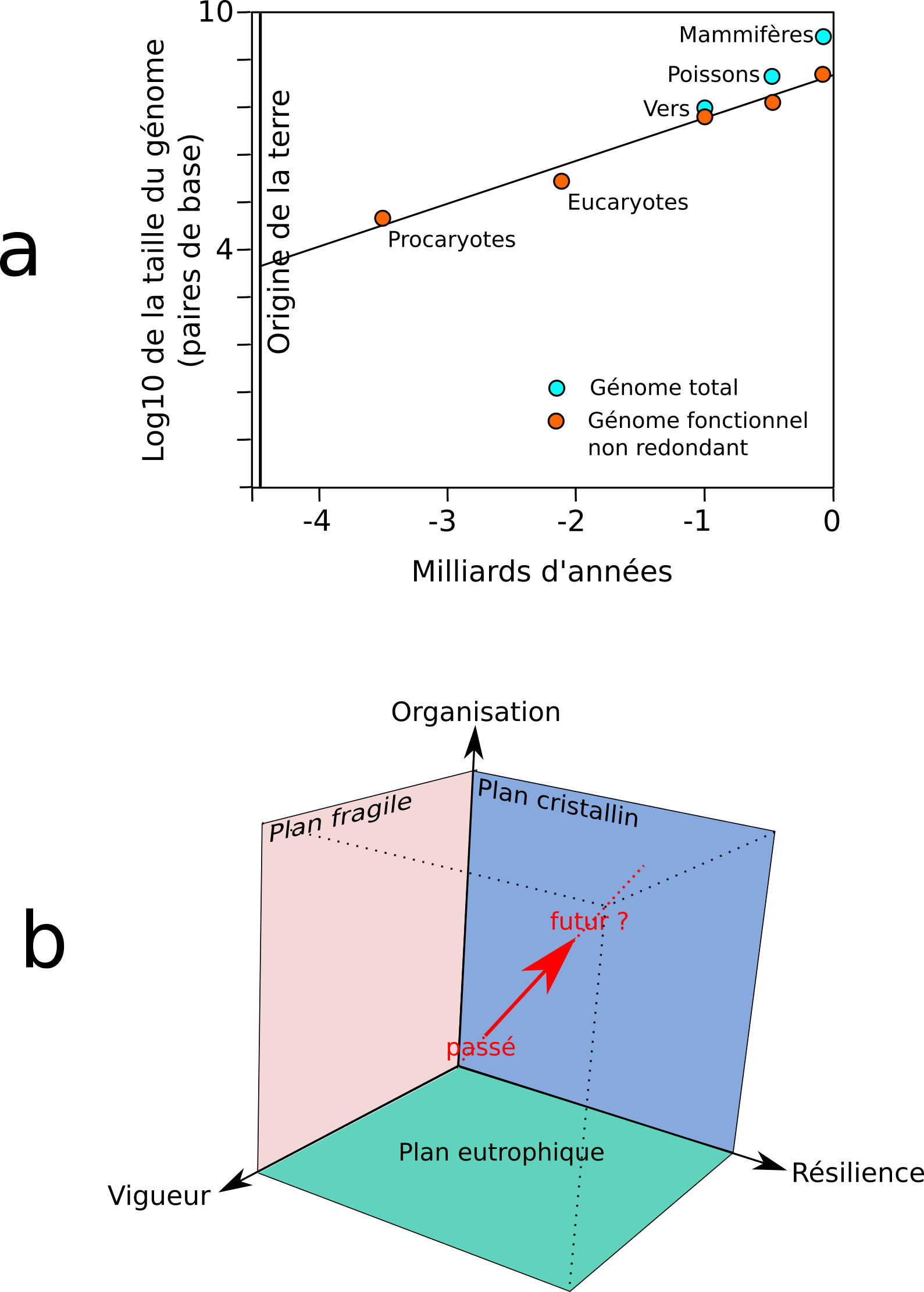
.
Figure 4 : Quel devenir pour la complexité des écosystèmes ?
a : selon Sharov et Gordon (2013), au cours de l’évolution, le génome des organismes suit une conjecture empirique de Moore (complexification croissante) ;
b : replacé dans l’espace tridimensionnel de Costanza (2012) l’organisation des écosystèmes devrait s’éloigner des 3 plans de cet espace (fragile, cristallin, eutrophique) où la valeur d’un des axes est nulle.
Conclusion
La question posée au chercheur en écologie comme au gestionnaire est donc de savoir s’il n’est d’autre choix que de réunir un consensus social autour des « services » attendus du système écologique considéré sachant que celui-ci, loin d’être un système « naturel » (au sens où Homo sapiens serait exclu) inclut l’activité humaine et en est même dans la plupart des paysages mondiaux depuis quelques milliers d’années le produit culturel sous forme de « socio-écosystème » (voir le regard n°4). Une question plus subtile, mais essentielle du point de vue du biologiste si l’on s’accorde sur la valeur intrinsèque de la biodiversité et les garanties qu’elle apporte sur la capacité des écosystèmes à évoluer, est de savoir quels scénarios sont à même d’assurer les meilleures capacités d’évolution.
Traduire cette question de gestion en un consensus social opérationnel reste à ce jour un défi pour le chercheur comme pour le gestionnaire, défi qui peut néanmoins s’appuyer sur divers courants de pensée portant sur l’éthique de la biodiversité (Maris, 2016) ou le principe de responsabilité des humains face au vivant proposé par Hans Jonas (Jonas, 1985). On comprend dès lors que les plans de gestion de la biodiversité puissent renvoyer à des concepts ambigus, quasi-fixistes,, assumant « à l’insu du plein gré » d’un opérateur qui connait pourtant les processus de l’évolution et la dynamique des écosystèmes une forme d’immuabilité des espèces et des écosystèmes actuels. Des zombies vivent encore parmi nous, et nous ne savons pas, conceptuellement et pratiquement, comment nous en débarrasser.
Bibliographie
Ceballos, G., Ehrlich, P.R., Barnosky, A.D., García, A., Pringle, R.M., Palmer, T.M., 2015. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Science Advances 1.
Chapin, F.S., Kofinas, G.P., Folke, C., 2009. Principles of Ecosystem Stewardship: Resilience-Based Natural Resource Management in a Changing World. Springer, New-York.
Costanza, R., 2012. Ecosystem health and ecological engineering. Ecological Engineering 45, 24-29.
Faith, D.P., Magallón, S., Hendry, A.P., Conti, E., Yahara, T., Donoghue, M.J., 2010. Evosystem services: an evolutionary perspective on the links between biodiversity and human well-being. Current Opinion in Environmental Sustainability 2, 66-74.
Giraudoux, P., 2014. Équilibre écologique et santé des écosystèmes: entre mythe biologique et consensus social. Nature ou Culture, Les colloques de l’Institut Universitaire de France. Publications de l’Université de St Etienne, Saint Etienne, pp. 129-142.
Jonas, H., 1985. Technik, Medizin und Ethik. Zur Praxis des Prinzips Verantwortung. Inselverlag, Frankfurt.
Mace, G.M., Masundire, H., Baillie, J.E.M., 2005. Biodiversity. Ecosystems and Human Well-Being: Current State and Trends. Island Press, Washington, DC, pp. 77–122.
Maris, V., 2016. Philosophie de la biodiversité: Petite éthique pour une nature en péril – Seconde édition. Buchet-Chastel, Paris.
Mayr, E., 1942. Mayr, Ernst (1942). Systematics and the Origin of Species. Columbia University Press., New York.
Millennium Ecosystem Assessment, 2005. Ecosystems and Human Well-being: Biodiversity Synthesis. World Resources Institute, Washington, DC.
Naeem, S., 1998. Species Redundancy and Ecosystem Reliability. Conserv. Biol. 12, 39-45.
Quigging, J., 2010. Zombie economics: how dead ideas still walk among us. Princeton University Press, Princeton.
Sarrazin, F., Lecomte, J., 2016. Evolution in the Anthropocene. Science 351, 922-923.
Teyssèdre, A., 2004. Vers une sixième grande crise d’extinctions? In: Barbault R. et Chevassus B. (Eds) et Teyssèdre A. (Coord.), Biodiversité et changements globaux : Enjeux de société et défis pour la recherche. ADPF, Paris, pp. 24-48.
—-
Regards connexes :
Barot S et F. Dubs: Les écosystèmes du sol. Regards et débats sur la biodiversité, SFE, Regard n°28, 17 février 2012.
Duru M., 2018. Agriculture, biodiversité et santé. Regards et débats sur la biodiversité, SFE2, Regard RO6, 12 sept. 2018.
Dussud C. et J-F. Ghiglione, 2014. La dégradation des plastiques en mer. Regards et débats sur la biodiversité, SFE, Regard n°63, 26 déc. 2014.
Faucon M-P., 2012. Ecologie et biodiversité des sites métallifères. Regards et débats sur la biodiversité, SFE, Regard n°32, mai 2012.
Julliard R. et M Burylo, 2012. Regard critique sur la compensation écologique. Regards et débats sur la biodiversité, SFE, Regard n°36, sept. 2012.
Lemoine G., 2013. La biodiversité des terrils. Regards et débats sur la biodiversité, SFE, Regard n°49, sept. 2013.
Mouquet N., I. Gounand et D. Gravelle, 2010. Biodiversité et fonctionnement des écosystèmes. Regards et débats sur la biodiversité, SFE, Regard n°3, 8 oct. 2010.
Pauly D. et F. Le Manach, 2012.Expansion et impacts de la pêche mondiale. Regards et débats sur la biodiversité, SFE, Regard n°31, 26 avril 2012.
Roche B. et A. Teyssèdre, 2011. Biodiversité et maladies infectieuses. Regards et débats sur la biodiversité, SFE, Regard n°18, 27 mai 2011.
Sarrazin et J. Lecomte, 2013. Peut-on dépasser l’antropomorphisme dans nos regards sur la biodiversité? Regards et débats sur la biodiversité, SFE, Regard n°59, juillet 2014.
Teyssèdre A., 2010. Les services écosystémiques, notion clé pour comprendre et préserver le fonctionnement des (socio)écosystèmes. Regards et débats sur la biodiversité, SFE, Regard n°4, 25 octobre 2010.
Thompson J. et O. Ronce. Fragmentation des habitats et dynamique de la biodiversité. Regards et débats sur la biodiversité, SFE, Regard n°6, 18 novembre 2010.
—
Article édité par A. Teyssèdre et S. Barot, mis en ligne par Anne Teyssèdre.
——
« Je soupçonne à présent que, de même qu’une harde de cerfs vit dans une peur mortelle du loup, la montagne vit dans une peur mortelle du cerf. Et avec plus de raison, parce qu’un cerf mâle pris par les loups sera remplacé en trois ans, mais un mont dénudé par les cerfs ne sera pas remplacé avant des décennies. De même avec les vaches. Car l’homme à bétail qui abat tous les loups de son ranch ne comprend pas qu’il doit alors assumer le rôle du loup, celui d’adapter le troupeau à la montagne. Il n’a pas appris à penser comme un montagne. D’où ces rivières qui arrachent l’avenir aux pentes, et le jettent à la mer. »
Aldo Leopold
Que savons nous, en réalité et à partir de nos préoccupations humaines, de ´la bonne échellé temporelle’ de modification des écosystèmes ?
Et quel est le ‘bon rapport’ entre la mesure d’un écosystème réel et la mesure de sa représentation utilitaire soustendant nos préoccupations anthropocentriques?